Яка будова та значення цитоплазми. Цитоплазма
Цитоплазма(cytoplasma) являє собою складну колоїдну систему, що складається з гіалоплазми, мембранних та немембранних органел та включень.
Гіалоплазма (від грец. hyaline - прозорий) є складною колоїдою системою, що складається з різних біополімерів (білки, нуклеїнові кислоти, полісахариди), яка здатна переходити із золеподібного (рідкого) стану в гель і назад.
Гіалоплазма складається з води, органічних та неорганічних сполук, розчинених у ній та цитоматриксу, представленого трабекулярною сіткою волокон білкової природи, товщиною 2-3 нм.
¨Функція гіалоплазми полягає в тому, що це середовище поєднує всі клітинні структури та забезпечує хімічну взаємодію їх один з одним.
Через гіалоплазму здійснюється більшість внутрішньоклітинних транспортних процесів: перенесення амінокислот, жирних кислот, нуклеотидів, цукрів. У гіалоплазмі йде постійний потік іонів до плазматичної мембрани та від неї, до мітохондрій, ядра та вакуолів. Гіалоплазма становить близько 50% всього об'єму цитоплазми.
Органели та включення. Органели - постійні та обов'язкові всім клітин мікроструктури, щоб забезпечити виконання життєво важливих функцій клітин.
Залежно від розмірів органели поділяються на:
1) мікроскопічні – видимі під світловим мікроскопом;
субмікроскопічні - помітні за допомогою електронного мікроскопа.
За наявності мембрани у складі органел розрізняють:
1) мембранні;
немембранні.
Залежно від призначення всі органели поділяються на:
Мембранні органели
Мітохондрії
Мітохондрії – мікроскопічні мембранні органели загального призначення.
Розміри - товщина 0,5мкм, довжина від 1 до 10мкм.
¨Форма - овальна, витягнута, неправильна.
Будова - мітохондрія обмежена двома мембранами товщиною близько 7нм:
1)Зовнішня гладка мітохондріальна мембрана(membrana mitochondrialis externa), яка відмежовує мітохондрію від гіалоплазми. Вона має рівні контури, замкнута таким чином, що представляє мішок.
Внутрішньою мітохондріальною мембраною(memrana mitochondrialis interna), яка утворює вирости, складки (кристи) всередину мітохондрії та обмежує внутрішній вміст мітохондрії – матрикс. Внутрішня частина мітохондрії заповнена електронно-щільною речовиною, яка має назву матрикс.
Матрикс має тонкозернисту будову і містить тонкі нитки завтовшки 2-3 нм та гранули розміром близько 15-20 нм. Нитки є молекулами ДНК, а дрібні гранули - мітохондріальні рибосоми.
¨Функції мітохондрій
1. Синтез і накопичення енергії у вигляді АТФ відбувається в результаті процесів окислення органічних субстратів і фосфорилювання АТФ. Ці реакції протікають за участю ферментів циклу трикарбонових кислот, локалізованих у матриксі. Мембрани кріст мають системи подальшого транспорту електронів та пов'язаного з ним окисного фосфорилювання (фосфорилювання АДФ до АТФ).
2. Синтез білка. Мітохондрії у своєму матриксі мають автономну систему синтезу білка. Це єдині органели, які мають молекули власної ДНК, вільної від гістонових білків. У матриксі мітохондрій також відбувається утворення рибосом, які синтезують ряд білків, що не кодуються ядром і використовуються для побудови власних ферментних систем.
3. Регулювання водного обміну.
Лізосоми
Лізосоми (lisosomae) – субмікроскопічні мембранні органели загального призначення.
Розміри - 0,2-0,4 мкм
¨Форма - овальна, дрібна, куляста.
Будова - лізосоми мають у своєму складі протеолітичні ферменти (відомо більше 60), які здатні розщеплювати різні біополімери. Ферменти розташовуються замкнутому мембранному мішечку, який запобігає їх попаданню в гіалоплазму.
Серед лізосом розрізняють чотири типи:
Первинні лізосоми;
Вторинні (гетерофагосоми, фаголізосоми);
Аутофагосоми
Залишкові тільця.
Первинні лізосоми- це дрібні мембранні бульбашки розміром 0,2-0,5 мкм, заповнені неструктурованою речовиною, що містить гідролітичні ферменти в неактивному стані (маркерно-кисла фосфотаза).
Вторинні лізосоми(гетерофагосоми) або внутрішньоклітинні травні вакуолі, які формуються при злитті первинних лізосом з фагоцитарними вакуолями. Ферменти первинної лізосоми починають контактувати з біополімерами і розщеплюють їх до мономерів. Останні транспортуються через мембрану в гіалоплазму, де відбувається їхня реутилізація, тобто включення в різні обмінні процеси.
Аутофагосоми (аутолізосоми)– постійно зустрічаються у клітинах найпростіших, рослин та тварин. За своєю морфологією їх відносять до вторинних лізосом, але з тією відмінністю, що у складі цих вакуолей зустрічаються фрагменти або навіть цілі цитоплазматичні структури, такі, як мітохондрії, пластиди, рибосоми, гранули глікогену.
Залишкові тільця(Телелізосома, corpusculum residuale) - являють собою оточені біологічною мембраною нерозщеплені залишки, містять невелику кількість гідролітичних ферментів, в них відбувається ущільнення вмісту, його перебудова. Часто в залишкових тільцях відбувається вторинна структуризація не перетравлених ліпідів і останні утворюють шаруваті структури. Там же спостерігається відкладення пігментних речовин – пігмент старіння, що містить ліпофусцин.
¨Функція - перетравлення біогенних макромолекул, модифікація продуктів, що синтезуються клітиною за допомогою гідролаз.
Цитоплазмаобов'язкова частина клітини, укладена між плазматичною мембраною і ядром і являє собою гіалоплазму -основна речовина цитоплазми, органоїди- постійні компоненти цитоплазми та включення- тимчасові компоненти цитоплазми. Хімічний склад цитоплазми різноманітний. Її основу становить вода (60-90% усієї маси цитоплазми). Цитоплазма багата на білки, до складу її можуть входити жири і жироподібні речовини, різні органічні та неорганічні сполуки. Цитоплазма має лужну реакцію. Одна з характерних рис цитоплазми - постійний рух (Циклоз).Воно виявляється, перш за все, переміщення органел клітини, наприклад хлоропластів. Якщо рух цитоплазми припиняється, клітина гине, оскільки, перебуваючи у постійному русі, може виконувати свої функції.
Основна речовина цитоплазми - гіалоплазма (цитозоль)- являє собою безбарвний, слизовий, густий та прозорий колоїдний розчин. Саме в ній протікають усі процеси обміну речовин, вона забезпечує взаємозв'язок ядра та всіх органоїдів. Залежно від переважання в гіалоплазмі рідкої частини або великих молекул розрізняють дві форми гіалоплазми: золь -більш рідка гіалоплазма та гель- густіша гіалоплазма. Між ними можливі взаємопереходи: гель легко перетворюється на золь і навпаки.
Клітинні оболонкиЕукаріотичні організми мають різну будову, але завжди до цитоплазми прилягає плазматична мембрана, на її поверхні утворюється зовнішній шар. У тварин він називається глікокаліксом(утворений глікопротеїнами, гліколіпідами, ліпопротеїнами), у рослин - клітинною стінкоюіз потужного шару волокон клітковини.
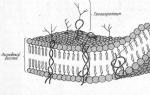
Будова мембран. Усі біологічні мембрани мають загальні структурні особливості та властивості. В даний час загальноприйнята рідинно-мозаїчна модельбудови мембрани (модель сендвіча). Основу мембрани становить ліпідний бислой, утворений в основному фосфоліпідами.У бислое хвости молекул у мембрані звернені друг до друга, а полярні головки - назовні, до води. Крім ліпідів у складі мембрани входять білки (в середньому 60%). Вони визначають більшість специфічних функцій мембрани. Молекули білків не утворюють суцільного шару, розрізняють периферичні білки- білки, що розташовуються на зовнішній або внутрішній поверхні ліпідного бислоя, напівінтегральні білки- білки, занурені в ліпідний бішар на різну глибину, інтегральні,або трансмембранні білки- білки, що пронизують мембрану наскрізь, контактуючи при цьому із зовнішньою, і з внутрішнім середовищем клітини.
Мембранні білки можуть виконувати різні функції: транспорт певних молекул, каталіз реакцій, що відбуваються на мембранах, підтримка структури мембран, отримання та перетворення сигналів з навколишнього середовища.
До складу мембран може входити від 2 до 10% вуглеводів. Вуглеводний компонент мембран зазвичай представлений олігосахаридними або полісахаридними ланцюгами, пов'язаними з молекулами білків (глікопротеїни) або ліпідів (гліколіпіди). В основному вуглеводи розміщуються на зовнішній поверхні мембрани. Вуглеводи забезпечують рецепторні функції мембрани. У тварин клітинах глікопротеїни утворюють надмембранний комплекс. глікоколікс,має товщину кілька десятків нанометрів. У ньому відбувається позаклітинне травлення, розташовуються багато рецепторів клітини, з його допомогою, мабуть, відбувається адгезія клітин.
Молекули білків і ліпідів рухливі, здатні переміщатися головним чином у площині мембрани. Товщина плазматичної мембрани загалом 7,5 нм.
Функції мембран.
1. Вони відокремлюють клітинний вміст від довкілля.
2. Регулюють обмін речовин між клітиною та середовищем.
3. Ділять клітини на компартаменти, призначені для перебігу різних реакцій.
4. Багато хімічних реакцій протікають на ферментативних конвеєрах, що розташовуються на самих мембранах.
5. Забезпечують зв'язок між клітинами у тканинах багатоклітинних організмів.
6. На мембранах розташовуються рецепторні ділянки для розпізнавання зовнішніх стимулів.
Одна з основних функцій мембрани - транспортна, що забезпечує обмін речовин між клітиною та зовнішнім середовищем. Мембрани мають властивість виборчої проникностітобто добре проникні для одних речовини або молекул і погано проникні (або зовсім непроникні) для інших. Існують різні механізми транспортування речовин через мембрану. Залежно від необхідності використання енергії для транспортування речовин розрізняють : пасивний транспорт- Транспорт речовин, що йде без витрат енергії; активний транспорт -транспорт, що йде із витратами енергії.
В основі пасивного транспорту лежить різниця концентрацій та зарядів. p align="justify"> При пасивному транспорті речовини завжди переміщуються з області з більш високою концентрацією в область з більш низькою, тобто по градієнту концентрації.
Розрізняють три основні механізми пасивного транспорту:проста дифузія- транспорт речовин безпосередньо через ліпідний бислой. Через нього легко проходять гази, неполярні чи малі незаряджені полярні молекули. Чим менше молекула і чим більше вона жиророзчинна, тим швидше вона проникає через мембрану. Цікаво, що полярні молекули води дуже швидко проникають через ліпідний бішар. Це тим, що її молекули малі і електрично нейтральні. Дифузію води через мембрани називають осмосом.
Дифузія через мембранні канали.Заряджені молекули та іони (Na + , К + , Са 2+ , С1~) не здатні проходити через ліпідний бислой шляхом простої дифузії, проте вони проникають через мембрану, завдяки наявності в ній особливих каналоутворюючих білків, що формують пори. Більшість води проходить мембрану через канали, утворені білками-аквапоринами.
Полегшена дифузія- транспорт речовин за допомогою спеціальних транспортних білків, кожен із яких відповідає за транспорт певних молекул або груп споріднених молекул. Вони взаємодіють з молекулою речовини, що переноситься, і яким-небудь способом переміщують її крізь мембрану. Так у клітину транспортуються цукру, амінокислоти, нуклеотиди та багато інших полярних молекул.
Необхідність активного транспортувиникає тоді, коли необхідно забезпечити перенесення через мембрану молекул проти електрохімічного градієнта. Цей транспорт здійснюється білками-переносниками, діяльність яких потребує витрат енергії. Джерелом енергії є молекули АТФ. Однією з найбільш вивчених систем активного транспорту є натрій-калієвий насос. Концентрація К+ усередині клітини значно вища, ніж за її межами, a Na+ – навпаки. Тому К+ через водяні пори мембрани пасивно дифундує з клітини, a Na+ – у клітину. Разом з тим для нормального функціонування клітини важливо підтримувати певне співвідношення іонів К+ та Na+ у цитоплазмі та зовнішньому середовищі. Це виявляється можливим тому, що мембрана завдяки наявності натрій-калієвого насоса активно перекачує Na + з клітини, а К + в клітину. На роботу натрій-калієвого насоса витрачається майже третина всієї енергії, яка потрібна для життєдіяльності клітини. За один цикл роботи насос викачує з клітини 3 іона Na + і закачує 2 іона К+. К + швидше пасивно дифундує з клітини, ніж Na + клітину.
Клітина має механізми, завдяки яким може здійснювати транспорт через мембрану великих частинок та макромолекул. Процес поглинання макромолекул клітиною називається ендоцитозом. При ендоцитозі плазматична мембрана утворює вп'ячування, краї її зливаються і відбувається відшнуровування в цитоплазму структур, відмежованих від цитоплазми одиночною мембраною, що є частиною зовнішньої цитоплазматичної мембрани. Розрізняють два типи ендоцитозу: фагоцитоз- захоплення та поглинання великих частинок (наприклад, фагоцитоз лімфоцитів, найпростіших та ін.) та піноцитоз -процес захоплення та поглинання крапельок рідини з розчиненими в ній речовинами.
Екзоцитоз- процес виведення різних речовин із клітини. При екзоцитозі мембрана бульбашки зливається із зовнішньою цитоплазматичною мембраною, вміст везикули виводиться за межі клітини, а її мембрана включається до складу зовнішньої цитоплазматичної мембрани.
Органоїди клітини
Органоїди (органели)- постійні клітинні структури, які забезпечують виконання клітиною специфічних функций. Кожен органоїд має певну будову та виконує певні функції.
Розрізняють: мембранні органоїди - мають мембранну будову, причому вони можуть бути одномембранними (ендоплазматичний ретикулум, апарат Гольджі, лізосоми, вакуолі рослинних клітин) та двомембранними (мітохондрії, пластиди, ядро).
Крім мембранних можуть бути і немембранні органоїди - не мають мембранної будови (хромосоми, рибосоми, клітинний центр і центріолі, вії та джгутики з базальними тільцями, мікротрубочки, мікрофіламенти).
Одномембранні органоїди:
1. Ендоплазматичний ретикулум (ЕПР).Є системою мембран, що формують цистерни і канали, з'єднаних один з одним і обмежують єдиний внутрішній простір - порожнини ЕПР.Мембрани з одного боку пов'язані із зовнішньою цитоплазматичною мембраною, з іншого - із зовнішньою оболонкою ядерної мембрани. Розрізняють два види ЕПР: шорсткий (гранулярний),містить рибосоми на своїй поверхні і являє собою сукупність сплощених мішечків, і гладкий (агранулярний),мембрани якого рибосом не несуть.
Функції: поділяє цитоплазму клітини на ізольовані відсіки, забезпечуючи тим самим просторове відмежування один від одного безлічі паралельних різних реакцій, Здійснює синтез і розщеплення вуглеводів і ліпідів (гладкий ЕПР) і забезпечує синтез білка (шорсткий ЕПР), накопичує потім транспортує до органоїдів клітини продукти біосинтезу.
2. Апарат Гольджі.Органоїд, зазвичай, розташований біля клітинного ядра (у тваринних клітинах часто поблизу клітинного центру). Є стопкою сплощених цистерн з розширеними краями, з якою пов'язана система дрібних одномембранних бульбашок (бульбашки Гольджі). Кожна стопка зазвичай складається з 4-6 цистерн. Число стосів Гольджі в клітці коливається від однієї до кількох сотень.
Найважливіша функція комплексу Гольджі - виведення з клітин різних секретів (ферментів, гормонів), тому він добре розвинений в секреторних клітинах. Тут відбувається синтез складних вуглеводів із простих цукрів, дозрівання білків, утворення лізосом.
3. Лізосоми.Найдрібніші одномембранні органоїди клітини, що являють собою бульбашки діаметром 0,2-0,8 мкм, містять до 60 гідролітичних ферментів, активних у слабокислому середовищі.
Освіта лізосом відбувається в апараті Гольджі, куди з ЕПР надходять синтезовані у ньому ферменти. Розщеплення речовин за допомогою ферментів називають лізисом, звідси і назва органоїду.
Розрізняють: первинні лізосоми - лізосоми, що відшнурувалися від апарату Гольджі та містять ферменти в неактивній формі, та вторинні лізосоми - лізосоми, що утворилися в результаті злиття первинних лізосом з піноцитозними або фагоцитозними вакуолями; у них відбувається перетравлення та лізис, що надійшли в клітину речовин (тому часто їх називають травними вакуолями).
Продукти перетравлення засвоюються цитоплазмою клітини, але частина матеріалу так і залишається неперетравленою. Вторинна лізосома, що містить цей неперетравлений матеріал, називається залишковим тільцем. Шляхом екзоцитозу неперетравлені частки видаляються із клітини.
Іноді за участю лізосом відбувається саморуйнування клітини. Цей процес називають автолізом. Зазвичай це відбувається при деяких процесах диференціювання (наприклад, заміна хрящової кісткової тканини, зникнення хвоста у пуголовка жаб).
4. Вії та джгутики.Утворені дев'ятьма здвоєними мікротрубочками, що утворюють стінку циліндра, покритого мембраною; у його центрі знаходяться дві одиночні мікротрубочки. Така структура типу 9+2 характерна для вій і джгутиків майже всіх еукаріотичних організмів, від найпростіших до людини.
Вії та джгутики укріплені в цитоплазмі базальними тільцями, що лежать на основі цих органоїдів. Кожне базальне тільце складається з дев'яти трійок мікротрубочок, у його центрі мікротрубочок немає.
5. До одномембранних органоїдів відносяться також і вакуолі, оточені мембраною - тонопластом. У рослинних клітинах можуть займати до 90% об'єму клітини та забезпечують надходження води в клітину за рахунок високого осмотичного потенціалу та тургор (внутрішньоклітинний тиск). У тварин клітинах вакуолі невеликі, утворюються за рахунок ендоцитозу (фагоцитозні та піноцитозні), після злиття з первинними лізосомами називаються травними вакуолями.
Двомембранні органоїди:
1. Мітохондрії. Двомембранні органоїди еукаріотичної клітини, які забезпечують організм енергією. Кількість мітохондрій у клітині коливається в широких межах, від 1 до 100 тис., та залежить від її метаболічної активності. Число мітохондрій може збільшуватися шляхом поділу, оскільки ці органоїди мають власну ДНК.
Зовнішня мембрана мітохондрій гладка, внутрішня мембрана утворює численні вп'ячування або трубчасті вирости. кристи. Число христ може коливатися від кількох десятків до кількох сотень і навіть тисяч, залежно від функцій клітини. Вони збільшують поверхню внутрішньої мембрани, де розміщуються ферментні системи, що у синтез молекул АТФ.
Внутрішній простір мітохондрій заповнений матриксом. У матриксі містяться кільцева молекула мітохондріальної ДНК специфічні іРНК, тРНК і рибосоми (прокаріотичного типу), що здійснюють автономний біосинтез частини білків, що входять до складу внутрішньої мембрани. Ці факти свідчать на користь походження мітохондрій від бактерій-окислювачів (згідно з гіпотезою симбіогенезу). Але більшість генів мітохондрії перейшла в ядро, і синтез багатьох мітохондріальних білків відбувається в цитоплазмі. Крім того, містяться ферменти, що утворюють молекули АТФ. Мітохондрії здатні розмножуватися шляхом розподілу.
Функції мітохондрій – кисневе розщеплення вуглеводів амінокислот, гліцерину та жирних кислот з утворенням АТФ, синтез мітохондріальних білків.
2. Пластиди. Розрізняють три основні типи пластид: лейкопласти- безбарвні пластиди в клітинах незабарвлених частин рослин, хромопласти- пофарбовані пластиди, зазвичай жовтого, червоного та оранжевого кольору, хлоропласти- Зелені пластиди. Пластиди утворюються з пропластиду - двомембранних бульбашок розміром до 1 мкм.
Оскільки пластиди мають загальне походження, між ними можливі взаємоперетворення. Найчастіше відбувається перетворення лейкопластів у хлоропласти (позеленіння бульб картоплі на світлі), зворотний процес відбувається в темряві. При пожовтінні листя та почервонінні плодів хлоропласти перетворюються на хромопласти. Вважають неможливим лише перетворення хромопластів на лейкопласти або хлоропласти.
Хлоропласти. Основна функція – фотосинтез, тобто. в хлоропластах світу здійснюється синтез органічних речовин з неорганічних за рахунок перетворення сонячної енергії в енергію молекул АТФ. Хлоропласти вищих рослин формою нагадують двоопуклу лінзу. Зовнішня мембрана гладка, а внутрішня має складчасту структуру. В результаті утворення випнувань внутрішньої мембрани виникає система ламелл і тилакоїдів. Внутрішнє середовище хлоропластів - стромамістить кільцеву ДНК та рибосоми прокаріотичного типу. Пластиди здатні до автономного поділу, як і мітохондрії. Факти, згідно з гіпотезою симбіогенезу, також свідчать на користь походження пластид від ціанобактерій.
Мал. Сучасна (узагальнена) схема будови рослинної клітини, Складена за даними електронно-мікроскопічного дослідження різних рослинних клітин: 1 - апарат Гольджі; 2 - вільно розташовані рибосоми; 3 – хлоропласти; 4 – міжклітинні простори; 5 - полірибосоми (кілька пов'язаних між собою рибосом); 6 - мітохондрії; 7 – лізосоми; 8 - гранульована ендоплазматична мережа; 9 – гладка ендоплазматична мережа; 10 - мікротрубочки; 11 – пластиди; 12 - плазмодесми, що проходять крізь оболонку; 13 – клітинна оболонка; 14 - ядерце; 15, 18 – ядерна оболонка; 16 - пори в ядерній оболонці; 17 - плазмалема; 19 – гіалоплазма; 20 - тонопласт; 21 - вакуолі; 22 – ядро.

Мал. Будова мембрани

Мал. Будова мітохондрії. Вгорі і в середині - вид поздовжнього зрізу через мітохондрію (вгорі - мітохондрія з ембріональної клітини кінчика кореня; у середині - з клітини дорослого листа елодеї). Внизу - тривимірна схема, де частина мітохондрії зрізана, що дозволяє бачити її внутрішню будову. 1 – зовнішня мембрана; 2 – внутрішня мембрана; 3 – кристи; 4 – матрикс.

Мал. Будова хлоропласту. Зліва - поздовжній розріз через хлоропласт: 1 - грани, утворені ламелами, складеними стосами; 2 – оболонка; 3 – строма (матрикс); 4 – ламели; 5 - краплі жиру, що утворився у хлоропласті. Справа - тривимірна схема розташування та взаємозв'язку ламелл і гран усередині хлоропласту: 1 - грани; 2 – ламели.
2. Будова цитоплазми, її хімічний склад, значення. Будова та функції мембран.
Цитоплазма (протоплазма)як живий вміст клітини відома була вже у XII столітті. Термін протоплазма вперше запропонований чеським ученим Пуркіньє (1839).
Розрізняють три шари цитоплазми: плазмалемму, гіалоплазму, тонопласт.
Плазмалема - Елементарна мембрана, зовнішній шар цитоплазми, примикає до оболонки. Товщина її близько 80А (А - ангстрем, 10-10 м). Складається із фосфоліпідів, білків, ліпопротеїнів, вуглеводів, неорганічних іонів, води. Може мати ламеллярну (шарувату) та міцелярну (краплинну) структури. Найчастіше складається з 3-х шарів: бимолекулярний шар фосфоліпідів (35А), на їхню частку припадає 40%, поверхня покрита з обох боків уривчастим шаром структурних білків (20 і 25А). У деяких місцях на стику ламеллярної та міцелярної структур або між двома міцелами зовнішній і внутрішній шари структурних білків можуть стулятися, утворюючи гідрофільні білкові пори, 7-10А, через які проходять речовини в розчиненому стані.
У матрикс мембран бувають вбудовані молекули білків, які мають ферментативної активності - специфічні селективні канали іонної провідності (калієві, натрієві та інших.). Нарешті, у мембрані може бути білки – ферменти, які забезпечують надходження у клітину високомолекулярних речовин. Всі ці утворення – біохімічні пори – забезпечують головну властивість мембран – напівпроникність.
Плазмалемма має численні складки, поглиблення, виступи, що збільшує її поверхню в багато разів.
Як мембрана, плазмалема виконує важливі та складні функції: 1. Регулює надходження та виділення речовин клітиною; 2. Перетворює, запасає та витрачає енергію; 3. Представляє хімічний перетворювач; прискорює перетворення речовин; 4. Приймає та перетворює світлові, механічні та хімічні сигнали зовнішнього світу.
Таким чином, плазмалема контролює проникність клітини, процеси поглинання, перетворення, секреції та екскреції речовин.
Тонопласт - Внутрішня мембрана, що відмежовує клітинний сік від цитоплазми
Гіалоплазма. Являє основу клітинної організації, є виразом її сутності як живого. З фізико-хімічної точки зору є складною гетерогенною колоїдною системою, де високомолекулярні сполуки дисперговані у водному середовищі. У середньому цитоплазма містить 70-80% води, 12% білків, 1,5-2% нуклеїнових кислот, близько 5% жиру, 4-6% вуглеводів та 0,5-2% неорганічних речовин. Може перебувати у двох станах: золю та гелю. Золь- рідкий стан, має в'язкість, гель- твердий стан, має еластичність, розтяжність. Здатна до оборотних переходів "золь-гель перехід" під впливом температури, концентрації водневих іонів, додавання електроліту, механічного впливу.
Цитоплазмазнаходиться в постійному русі, яке у звичайних умовах дуже повільне та майже непомітне. Підвищення температури, світловий чи хімічний подразник прискорюють рух цитоплазми і роблять його помітним у світловому мікроскопі. Побачити цей рух допомагають хлоропласти, які захоплюються струмом в'язкої цитоплазми. Рух цитоплазми буває двох видів: круговий (ротаційний) та струменевий (циркуляційний). Якщо порожнина клітини зайнята однією великою вакуоллю, то цитоплазма рухається лише вздовж стінок. Це круговий рух. Його можна спостерігати в клітинах листа валліснерії, елодеї. Якщо у клітині кілька вакуолей, то тяжі цитоплазми, перетинаючи клітину, з'єднуються у центрі, де розташовується ядро. У цих тяжах відбувається струйчастий рух цитоплазми. Струменевий рух цитоплазми можна спостерігати в клітинах пекучих волосків кропиви, в клітинах волосків молодих пагонів гарбуза.
Властивості гіалоплазми пов'язані з надмолекулярними структурами білкової природи. Це мікротрубочки та мікрофіламенти.
Мікротрубочки- порожнисті дрібні утворення з електроноплотною білковою стінкою. Беруть участь у проведенні речовин по цитоплазмі, у переміщенні хромосом та утворенні ниток мітотичного веретена.
Мікрофіламентискладаються із спірально розташованих білкових субодиниць, що утворюють волокна або тривимірну мережу, містять скорочувальні білки та сприяють руху гіалоплазми та прикріплених до них органоїдів.
Гіалоплазма як складна гетерогенна колоїдна система макромолекул і надмолекулярних структур характеризується нерозчинністю у воді, в'язкістю, еластичністю, здатністю до зворотних змін, непрохідністю через пори природних мембран, великими поверхнями розділу, має сильне світлозаломлення, дуже малу швидкість дифузії.
Органоїди гіалоплазми . Як зазначалося раніше, у гіалоплазмі є велика кількість надмолекулярних утворень, які являють собою численні органоїди.
Функції біомембран
1) бар'єрна - забезпечує регульований, вибірковий, пасивний та активний обмін речовин з навколишнім середовищем. Наприклад, мембрана пероксисом захищає цитоплазму від небезпечних для клітин пероксидів. Виборча проникність означає, що проникність мембрани для різних атомів або молекул залежить від їх розмірів, електричного заряду та хімічних властивостей. Виборча проникність забезпечує відокремлення клітини та клітинних компартментів від навколишнього середовища та постачання їх необхідними речовинами.
2)транспортна - через мембрану відбувається транспорт речовин у клітину та з клітини. Транспорт через мембрани забезпечує: доставку поживних речовин, видалення кінцевих продуктів обміну, секрецію різних речовин, створення іонних градієнтів, підтримання в клітині відповідного pH та іонної концентрації, які потрібні для роботи клітинних ферментів. (наприклад, через гідрофільні властивості, оскільки мембрана всередині гідрофобна і не пропускає гідрофільні речовини, або через великі розміри), але необхідні для клітини, можуть проникнути крізь мембрану через спеціальні білки-переносники (транспортери) і білки-канали або шляхом ендоцитозу. При пасивному транспорті речовини перетинають ліпідний бислой без витрат енергії шляхом дифузії. Варіантом цього механізму є полегшена дифузія, за якої речовині допомагає пройти через мембрану будь-яка специфічна молекула. У цієї молекули може бути канал, що пропускає речовини тільки одного типу. Активний транспорт вимагає витрат енергії, оскільки відбувається проти концентрації градієнта. На мембрані існують спеціальні білки-насоси, у тому числі АТФаза, яка активно вкачують у клітину іони калію (K+) та викачують із неї іони натрію (Na+).
3) матрична - забезпечує певне взаєморозташування та орієнтацію мембранних білків, їх оптимальну взаємодію;
4)механическая - забезпечує автономність клітини, її внутрішньоклітинних структур, і навіть з'єднання коїться з іншими клітинами (в тканинах). Велику роль забезпечення механічної функції мають клітинні стінки, а тварин - міжклітинна речовина.
5) енергетична - при фотосинтезі в хлоропластах та клітинному диханні в мітохондріях у їх мембранах діють системи перенесення енергії, в яких також беруть участь білки;
6) рецепторна - деякі білки, що знаходяться в мембрані, є рецепторами (молекулами, за допомогою яких клітина сприймає ті чи інші сигнали). Наприклад, гормони, що циркулюють у крові, діють тільки на такі клітини-мішені, у яких є відповідні цим гормонам рецептори . Нейромедіатори (хімічні речовини, що забезпечують проведення нервових імпульсів) теж зв'язуються з особливими білками рецепторів клітин-мішеней.
7) ферментативна – мембранні білки нерідко є ферментами. Наприклад, плазматичні мембрани епітеліальних клітин кишківника містять травні ферменти.
8) здійснення генерації та проведення біопотенціалів.
За допомогою мембрани в клітині підтримується постійна концентрація іонів: концентрація іону К+ усередині клітини значно вища, ніж зовні, а концентрація Na+ значно нижча, що дуже важливо, оскільки це забезпечує підтримку різниці потенціалів на мембрані та генерацію нервового імпульсу.
9) маркування клітини – на мембрані є антигени, що діють як маркери – «ярлики», що дозволяють пізнати клітину. Це глікопротеїни (тобто білки з приєднаними до них розгалуженими олігосахаридними бічними ланцюгами), що відіграють роль «антен». Через незліченну безліч конфігурації бічних ланцюгів можна створити для кожного типу клітин свій спеціальний маркер. За допомогою маркерів клітини можуть розпізнавати інші клітини і діяти відповідно до них, наприклад, при формуванні органів і тканин. Це дозволяє імунній системі розпізнавати чужорідні антигени.
Гелеподібний вміст клітини, обмежений мембраною називається цитоплазмою живої клітини. Поняття було запроваджено 1882 року німецьким ботаніком Едуардом Страсбургером.
Будова
Цитоплазма є внутрішнім середовищем будь-якої клітини і характерна для клітин бактерій, рослин, грибів, тварин.
Цитоплазма складається з наступних компонентів:
- гіалоплазми (цитозолі) - рідкої речовини;
- клітинних включень – необов'язкових компонентів клітини;
- органоїдів – постійних компонентів клітини;
- цитоскелета – клітинного каркаса.
Хімічний склад цитозолі містить такі речовини:
- воду – 85 %;
- білки - 10%
- органічні сполуки – 5 %.
До органічних сполук відносяться:
- мінеральні солі;
- вуглеводи;
- ліпіди;
- азотовмісні сполуки;
- незначна кількість ДНК та РНК;
- глікоген (характерний для тварин клітин).

Мал. 1. Склад цитоплазми.
Цитоплазма містить запас поживних речовин (краплі жиру, зерна полісахаридів), а також нерозчинні відходи життєдіяльності клітини.
Цитоплазма безбарвна та постійно рухається, перетікає. Вона містить усі органели клітини та здійснює їх взаємозв'язок. При частковому видаленні цитоплазма відновлюється. При повному видаленні цитоплазми клітка гине.
Будова цитоплазми неоднорідна. Умовно виділяють два шари цитоплазми:
ТОП-4 статтіякі читають разом з цією
- ектоплазму (плазмагель) - зовнішній щільний шар, що не містить органел;
- ендоплазму (плазмазоль) - внутрішній рідкіший шар, що містить органели.
Поділ на ектоплазму та ендоплазму яскраво виражений у найпростіших. Ектоплазма допомагає клітині пересуватися.
Зовні цитоплазма оточена цитоплазматичною мембраною чи плазмалемою. Вона захищає клітину від пошкоджень, здійснює вибірковий транспорт речовин та забезпечує подразливість клітини. Мембрана складається з ліпідів та білків.
Життєдіяльність
Цитоплазма - життєво важлива речовина, яка бере участь у головних процесах клітини:
- метаболізмі;
- зростанні;
- розподілі.
Рух цитоплазми називається циклозом чи цитоплазматичним потоком. Він здійснюється у клітинах еукаріотів, у тому числі й людини. При циклозі цитоплазма доставляє речовини всім органел клітини, здійснюючи клітинний метаболізм. Переміщається цитоплазма у вигляді цитоскелета з витратою АТФ.
У разі збільшення обсягу цитоплазми клітина зростає. Процес поділу тіла еукаріотичної клітини після поділу ядра (каріокінезу) називається цитокінезом. Внаслідок поділу тіла цитоплазма разом із органелами розподіляється між двома дочірніми клітинами.

Мал. 2. Цитокінез.
Функції
Основні функції цитоплазми у клітині описані у таблиці.
Відділення цитоплазми від мембрани при осмосі води, що виходить назовні, називається плазмолізом. Зворотний процес - деплазмоліз - відбувається при надходженні до клітини достатньої кількості води. Процеси притаманні будь-яких клітин, крім тваринної.
Цитоплазма- обов'язкова частина клітини, укладена між плазматичною мембраною та ядром; підрозділяється на гіалоплазму (основна речовина цитоплазми), органоїди (постійні компоненти цитоплазми) та включення (тимчасові компоненти цитоплазми). Хімічний склад цитоплазми: основу становить вода (60-90% усієї маси цитоплазми), різні органічні та неорганічні сполуки. Цитоплазма має лужну реакцію. Характерна риса цитоплазми еукаріотичної клітини - постійний рух ( циклоз). Воно виявляється, перш за все, переміщення органоїдів клітини, наприклад хлоропластів. Якщо рух цитоплазми припиняється, клітина гине, оскільки, перебуваючи у постійному русі, може виконувати свої функції.
Гіалоплазма ( цитозоль) являє собою безбарвний, слизовий, густий та прозорий колоїдний розчин. Саме в ній протікають усі процеси обміну речовин, вона забезпечує взаємозв'язок ядра та всіх органоїдів. Залежно від переважання в гіалоплазмі рідкої частини або великих молекул розрізняють дві форми гіалоплазми: золь- Рідкіша гіалоплазма і гель- Густіша гіалоплазма. Між ними можливі взаємопереходи: гель перетворюється на золь і навпаки.
Функції цитоплазми:
- поєднання всіх компонентів клітини в єдину систему,
- середовище для проходження багатьох біохімічних та фізіологічних процесів,
- середовище для існування та функціонування органоїдів.
Клітинні оболонки
Клітинні оболонкиобмежують еукаріотичні клітини. У кожній клітинній оболонці можна виділити щонайменше два шари. Внутрішній шар прилягає до цитоплазми та представлений плазматичною мембраною(синоніми - плазмалема, клітинна мембрана, цитоплазматична мембрана), над якою формується зовнішній шар. У тваринній клітці він тонкий і називається глікокаліксом(утворений глікопротеїнами, гліколіпідами, ліпопротеїнами), у рослинній клітині - товстий, називається клітинною стінкою(Утворений целюлозою).
Усі біологічні мембрани мають загальні структурні особливості та властивості. В даний час загальноприйнята рідинно-мозаїчна модель будови мембрани. Основу мембрани становить ліпідний бислой, утворений переважно фосфоліпідами. Фосфоліпіди - тригліцериди, у яких один залишок жирної кислоти заміщений залишок фосфорної кислоти; Ділянку молекули, в якій знаходиться залишок фосфорної кислоти, називають гідрофільною головкою, ділянки, в яких знаходяться залишки жирних кислот - гідрофобними хвостами. У мембрані фосфоліпіди розташовуються строго впорядковано: гідрофобні хвости молекул звернені одна до одної, а гідрофільні головки - назовні, до води.
Крім ліпідів до складу мембрани входять білки (у середньому 60%). Вони визначають більшість специфічних функцій мембрани (транспорт певних молекул, каталіз реакцій, отримання та перетворення сигналів із навколишнього середовища та ін.). Розрізняють: 1) периферичні білки(розташовані на зовнішній або внутрішній поверхні ліпідного бислоя), 2) напівінтегральні білки(занурені в ліпідний бішар на різну глибину), 3) інтегральні, або трансмембранні, білки(Пронизують мембрану наскрізь, контактуючи при цьому і із зовнішньою, і з внутрішнім середовищем клітини). Інтегральні білки в ряді випадків називають каналоутворюючими, або канальними, так як їх можна розглядати як гідрофільні канали, якими в клітину проходять полярні молекули (ліпідний компонент мембрани їх би не пропустив).

А - гідрофільна головка фосфоліпіду; В – гідрофобні хвостики фосфоліпіду; 1 - гідрофобні ділянки білків Е та F; 2 - гідрофільні ділянки білка F; 3 - розгалужена олігосахаридна ланцюг, приєднана до ліпіду в молекулі гліколіпіду (гліколіпіди зустрічаються рідше, ніж глікопротеїни); 4 - розгалужена олігосахаридна ланцюг, приєднана до білка в молекулі глікопротеїну; 5 - гідрофільний канал (функціонує як пора, через яку можуть проходити іони та деякі полярні молекули).
До складу мембрани можуть входити вуглеводи (до 10%). Вуглеводний компонент мембран представлений олігосахаридними або полісахаридними ланцюгами, пов'язаними з молекулами білків (глікопротеїни) або ліпідів (гліколіпіди). В основному вуглеводи розміщуються на зовнішній поверхні мембрани. Вуглеводи забезпечують рецепторні функції мембрани. У тваринних клітинах глікопротеїни утворюють надмембранний комплекс - глікокалікс, що має товщину кілька десятків нанометрів. У ньому розташовуються багато рецепторів клітини, з його допомогою відбувається адгезія клітин.
Молекули білків, вуглеводів та ліпідів рухливі, здатні переміщатися у площині мембрани. Товщина плазматичної мембрани – приблизно 7,5 нм.
Функції мембран
Мембрани виконують такі функції:
- відділення клітинного вмісту від зовнішнього середовища,
- регуляція обміну речовин між клітиною та середовищем,
- поділ клітини на компартаменти («відсіки»),
- місце локалізації «ферментативних конвеєрів»,
- забезпечення зв'язку між клітинами у тканинах багатоклітинних організмів (адгезія),
- розпізнавання сигналів.
Найважливіше властивість мембран- Виборча проникність, тобто. мембрани добре проникні одних речовин чи молекул і погано проникні (чи зовсім непроникні) іншим. Ця властивість лежить в основі регуляторної функції мембран, що забезпечує обмін речовин між клітиною та зовнішнім середовищем. Процес проходження речовин через клітинну мембрану називають транспортом речовин. Розрізняють: 1) пасивний транспорт- Процес проходження речовин, що йде без витрат енергії; 2) активний транспорт- Процес проходження речовин, що йде з витратами енергії.
При пасивному транспортіречовини переміщаються з області з більш високою концентрацією область з нижчою, тобто. за градієнтом концентрації. У будь-якому розчині є молекули розчинника та розчиненої речовини. Процес переміщення молекул розчиненої речовини називають дифузією, переміщення молекул розчинника осмосом. Якщо молекула заряджена, то її транспорт впливає і електричний градієнт. Тому часто говорять про електрохімічний градієнт, об'єднуючи обидва градієнти разом. Швидкість транспорту залежить від величини градієнта.
Можна виділити такі види пасивного транспорту: 1) проста дифузія- Транспорт речовин безпосередньо через ліпідний бішар (кисень, вуглекислий газ); 2) дифузія через мембранні канали- Транспорт через каналоутворюючі білки (Na +, K +, Ca 2+, Cl -); 3) полегшена дифузіятранспорт речовин за допомогою спеціальних транспортних білків, кожен з яких відповідає за переміщення певних молекул або груп споріднених молекул (глюкоза, амінокислоти, нуклеотиди); 4) осмос- Транспорт молекул води (у всіх біологічних системах розчинником є саме вода).
Необхідність активного транспортувиникає тоді, коли необхідно забезпечити перенесення через мембрану молекул проти електрохімічного градієнта. Цей транспорт здійснюється особливими білками-переносниками, діяльність яких потребує витрат енергії. Джерелом енергії є молекули АТФ. До активного транспорту відносять: 1) Na+/К+-насос (натрій-калієвий насос); 2) ендоцитоз; 3) екзоцитоз.

Робота Na+/К+-насоса. Для нормального функціонування клітина повинна підтримувати певне співвідношення іонів К+ та Na+ у цитоплазмі та у зовнішньому середовищі. Концентрація К + усередині клітини має бути значно вищою, ніж за її межами, а Na + - навпаки. Слід зазначити, що Na + і + можуть вільно дифундувати через мембранні пори. Na + /К + -насос протидіє вирівнюванню концентрацій цих іонів і активно перекачує Na + з клітини, а K + в клітину. Na + /К + -насос є трансмембранний білок, здатний до конформаційних змін, внаслідок чого він може приєднувати як K + , так і Na + . Цикл роботи Na + /К + -насоса можна розділити на наступні фази: 1) приєднання Na + з внутрішньої сторони мембрани; 2) фосфорилювання білка-насоса; 3) вивільнення Na + у позаклітинному просторі; 4) приєднання K + із зовнішньої сторони мембрани. 5) дефосфорилювання білка-насоса; 6) вивільнення K+ у внутрішньоклітинному просторі. На роботу натрій-калієвого насоса витрачається майже третина всієї енергії, яка потрібна для життєдіяльності клітини. За один цикл роботи насос викачує із клітини 3Na+ та закачує 2К+.
Ендоцитоз- Процес поглинання клітиною великих частинок та макромолекул. Розрізняють два типи ендоцитозу: 1) фагоцитоз- Захоплення та поглинання великих частинок (клітин, частин клітин, макромолекул) і 2) піноцитоз- Захоплення та поглинання рідкого матеріалу (розчин, колоїдний розчин, суспензія). Явище фагоцитозу відкрито І.І. Мечниковим у 1882 р. При ендоцитозі плазматична мембрана утворює вп'ячування, краї її зливаються, і відбувається відшнурування в цитоплазму структур, відмежованих від цитоплазми одиночною мембраною. До фагоцитозу здатні багато найпростіших, деякі лейкоцити. Піноцитоз спостерігається в епітеліальних клітинах кишківника, в ендотелії кровоносних капілярів.
Екзоцитоз- Процес, зворотний ендоцитоз: виведення різних речовин з клітини. При екзоцитозі мембрана бульбашки зливається із зовнішньою цитоплазматичною мембраною, вміст везикули виводиться за межі клітини, а її мембрана включається до складу зовнішньої цитоплазматичної мембрани. У такий спосіб із клітин залоз внутрішньої секреції виводяться гормони, у найпростіших — неперетравлені залишки їжі.
Перейти до лекції №5«Клітинна теорія. Типи клітинної організації»
Перейти до лекції №7«Еукаріотична клітина: будова та функції органоїдів»