Хто сформулював закон зародкової схожості. Співвідношення онтофілогенезу
Закон зародкової подібності ембріологія тварин
Загальна ембріологія: Термінологічний словник.. О.В. Ділекова, Т.І. Лапіна. 2010 .
Дивитись що таке "закон зародкової схожості" в інших словниках:
ЕМБРІОЛОГІЯ ТВАРИННИХ ЗАКОН БЕРА, ЗАКОН ЗАРОДИШОВОГО подібності – закон був сформульований Карлом Ернестом фон Бером у 1828 році: Загальні основні риси, характерні для будь-якої великої групи тварин, виявляються в процесі розвитку раніше, ніж… Загальна ембріологія: Термінологічний словник
Біогенетичний закон- Геккеля Мюллера (також відомий під назвами «закон Геккеля», «закон Мюллера Геккеля», «закон Дарвіна Мюллера Геккеля», «основний біогенетичний закон»): кожна жива істота у своєму індивідуальному розвитку (онтогенез) повторює у відомій…
БІОГЕНЕТИЧНИЙ ЗАКОН- (від грец. bios – життя і genesis – походження) вперше сформульоване Фріцем Мюллером та Ернстом Геккелем положення про те, що ряд форм, які жива істота проходить у процесі свого індивідуального розвитку від яйцеклітини до розвиненого… Філософська енциклопедія
Порівняльна анатомія тварин- Порівняльна морфологія, наука, що вивчає закономірності будови та розвитку органів та їх систем шляхом зіставлення у тварин різних систематичних груп. Порівняння будови органів у зв'язку з їх функціями дає можливість зрозуміти… Велика Радянська Енциклопедія
ПОРІВНЯЛЬНА АНАТОМІЯ ТВАРИН- Порівняльна морфологія, розділ морфології тварин, що вивчає закономірності будови та розвитку органів та їх систем шляхом зіставлення тварин різних систематич. груп. Порівняння будови органів у зв'язку з їх функціями дає можливість… Біологічний енциклопедичний словник
Докази еволюції- Викопний археоптерикс, виявлений невдовзі після публікації «… Вікіпедія
Макроеволюція- органічного світу це процес формування великих систематичних одиниць: з видів нових пологів, з пологів нових сімейств і т. д. В основі макроеволюції лежать ті ж рушійні сили, що і в основі мікроеволюції:
ембріологія- та; ж. [від грец. embryon зародок і logos вчення] Розділ біології, що вивчає освіту та розвиток ембріонів. е. тварин. Порівняльна е. ◁ Ембріологічний, ая, ое. Це дослідження. * * * ембріологія (від ембріон і...логія), наука про… Енциклопедичний словник
Еволюційне вчення- (також еволюціонізм та еволюціоністика) система ідей та концепцій у біології, що стверджують історичний прогресивний розвиток біосфери Землі, що становлять її біогеоценозів, а також окремих таксонів та видів, яке може бути вписано в… … Вікіпедія
Історія еволюційного вчення- бере початок у античних філософських системах, ідеї яких, своєю чергою, коренилися у космологічних міфах. Поштовхом до визнання еволюції науковою спільнотою стала публікація книги Чарльза Дарвіна «Походження видів шляхом природного… … Вікіпедія
Онтогенез - Реалізація генетичної інформації, що відбувається на всіх стадіях.
Онтогенез – генетично контрольований процес. У ході онтогенезу реалізується генотип та формується фенотип.
Онтогенез – індивідуальний розвиток організму, сукупність послідовних морфологічних, фізіологічних та біохімічних перетворень, що зазнають організму від моменту його зародження до кінця життя. О. включає зростання, тобто збільшення маси тіла, його розмірів, диференціювання. Термін "О." введений Е. Геккелем (1866) при формулюванні ним біогенетичного закону.
Першу спробу історичного обґрунтування О. зробив І. ф. Меккель. Проблема співвідношення О. та філогенезу була поставлена Ч. Дарвіном і розроблялася Ф. Мюллером,Е. Всі пов'язані зі зміною спадковості, нові в еволюційному відношенні ознаки виникають в О., але лише ті з них, які сприяють кращому пристосуванню організму до умов існування, зберігаються в процесі природного відбору і передаються наступним поколінням, тобто закріплюються у еволюції. p align="justify"> Пізнання закономірностей, причин і факторів О. служить науковою основою для відшукання засобів впливу на розвиток рослин, тварин і людини, що має найважливіше значення для практики рослинництва і тваринництва, а також для медицини.
Філогенез – історичний розвиток організмів. Термін введений в ньому. Еволюціоніст Е. Геккель в 1866. Основним завданням при вивченні Ф. є реконструкція еволюційних перетворень тварин, рослин, мікроорганізмів, встановлення на цій основі їх походження і споріднених зв'язків між таксонами, до яких належать вивчені організми. Для цієї мети Е. Геккель розробив метод "потрійного паралелізму", що дозволяє шляхом зіставлення даних трьох наук - морфології, ембріології та палеонтології - відновити хід історичного розвитку систематичної групи, що вивчається.
Закон зародкової подібності
Дослідники початку ХІХ ст. вперше почали звертати увагу на подібність стадій розвитку ембріонів вищих тварин із ступенями ускладнення організації, що ведуть від низькоорганізованих форм до прогресивних. Порівнюючи стадії розвитку зародків різних видів та класів хордових, К. Бер зробив такі висновки.
1. Ембріони тварин одного типу на ранніх стадіях розвитку подібні.
2. Вони послідовно переходять у своєму розвитку від більш загальних ознак типу до більш приватних. В останню чергу розвиваються ознаки, що вказують на приналежність ембріона до певного роду, виду і, нарешті, індивідуальні риси.
3. Ембріони різних представників одного типу поступово відокремлюються один від одного.
К. Бер, будучи еволюціоністом, було пов'язувати відкриті їм закономірності індивідуального розвитку з процесом филогенеза. Тому зроблені ним узагальнення мали значення лише емпіричних правил.
Розвиток еволюційної ідеї надалі дозволило пояснити подібність ранніх зародків їх історичним спорідненістю, а придбання ними дедалі більше приватних рис із поступовим відокремленням друг від друга - дійсним відокремленням відповідних класів, загонів, сімейств, пологів та видів у процесі еволюції.
Незабаром після відкриття закону зародкової подібності Ч. Дарвін показав, що цей закон свідчить про спільність походження та єдність початкових етапів еволюції в межах типу.
Біогенетичний законГеккеля-Мюллера: кожна жива істота у своєму індивідуальному розвитку (онтогенез) повторює певною мірою форми, пройденого його предками або його видом (філогенез).
Онтогенез – повторення філогенезу
Зіставляючи онтогенез ракоподібних з морфологією їх вимерлих предків, Ф. Мюллер зробив висновок про те, що ракоподібні, що нині живуть, у своєму розвитку повторюють шлях, пройдений їх предками. Перетворення онтогенезу на еволюції, на думку Ф. Мюллера, здійснюється завдяки його подовженню з допомогою додавання до нього додаткових стадій чи надставок. На основі цих спостережень, а також вивчення розвитку хордових Е. Геккель (1866) сформулював основний біогенетичний закон, відповідно до якого онтогенез є коротким і швидким повторенням філогенезу.
Повторення структур, притаманних предків, в ембріогенезі нащадків названо рекапітуляціями.Рекапітулують не тільки морфологічні ознаки - хорда, закладки зябрових щілин і зябрових дуг у всіх хордових, а й особливості біохімічної організації та фізіології. Так було в еволюції хребетних відбувається поступова втрата ферментів, необхідні розпаду сечової кислоти - продукту метаболізму пуринів. У більшості безхребетних кінцевий продукт розпаду сечової кислоти - аміак, у земноводних і риб - сечовина, у багатьох плазунів - алантоїн, а у деяких ссавців сечова кислота взагалі не розщеплюється і виділяється із сечею. В ембріогенезі ссавців та людини відзначені біохімічні та фізіологічні рекапітуляції: виділення ранніми зародками аміаку, пізніше сечовини, потім алантоїну, а на останніх стадіях розвитку – сечової кислоти.
Однак у онтогенезі високоорганізованих організмів який завжди спостерігається суворе повторення стадій історичного поступу, як це випливає з біогенетичного закону. Так, зародок людини ніколи не повторює дорослих стадій риб, земноводних, плазунів і ссавців, а подібний до ряду рис лише з їхніми зародками. Ранні стадії розвитку зберігають найбільшу консервативність, завдяки чому рекапітулують повніше, ніж пізні. Це пов'язано з тим, що одним з найбільш важливих механізмів інтеграції ранніх етапів ембріогенезу є ембріональна індукція, а структури зародка, що формуються в першу чергу, такі, як хорда, нервова трубка, ковтка, кишка та соміти, є організаційними центрами зародка, від яких залежить весь перебіг розвитку.
Генетична основа рекапітуляції полягає в єдності механізмів генетичного контролю розвитку, що зберігається на базі загальних генів регуляції онтогенезу, які дістаються родинним групам організмів від спільних предків.
Рекапітуляція(від латів. recapitulatio – повторення) - поняття, що використовується в біології для позначення повторення в індивідуальному розвитку ознак, властивих ранній стадії еволюційного розвитку.
Квиток 96.
Онтогенез як основа філогенезу. Ціногенези. Автономізація онтогенезу. Філембріогенези. Вчення А.Н.Северцова про філембріогенези. Механізми їхнього виникнення. Гетерохронії та гетеротопії біологічних структур в еволюції онтогенезу.
Спираючись лише на основний біогенетичний закон, неможливо пояснити процес еволюції: нескінченне повторення пройденого саме собою не народжує нового. Так як життя існує на Землі завдяки зміні поколінь конкретних організмів, еволюція її протікає завдяки змінам, що відбуваються в їх онтогенезі. Ці зміни зводяться до того, що конкретні онтогенези відхиляються від шляху, прокладеного предковими формами, і набувають нових рис.
До таких відхилень відносяться, наприклад, ціногенези -пристосування, що виникають у зародків або личинок і адаптують їх до особливостей довкілля. У дорослих організмів ціногенези не зберігаються. Прикладами ценогенезів є рогові утворення у роті личинок безхвостих земноводних, що полегшують їм живлення рослинною їжею. У процесі метаморфозу у жабки вони зникають і травна система перебудовується для живлення комахами та хробаками. До ценогенезів у амніот відносять зародкові оболонки, жовтковий мішок та алантоїс, а у плацентарних ссавців та людини – ще й плаценту з пуповиною.
Ціногенези, виявляючись лише на ранніх стадіях онтогенезу, не змінюють типу організації дорослого організму, але забезпечують більш високу ймовірність виживання потомства. Вони можуть супроводжуватися при цьому зменшенням плодючості та подовженням зародкового або личинкового періоду, завдяки чому організм у постембріональному або післяличинковому періоді розвитку виявляється більш зрілим та активним. Виникнувши та виявившись корисними, ценогенези відтворюватимуться у наступних поколіннях. Так, амніон, що з'явився вперше у предків плазунів у кам'яновугільному періоді палеозойської ери, відтворюється у всіх хребетних, що розвиваються на суші, як у яйцекладних - плазунів і птахів, так і у плацентарних ссавців.
Інший тип філогенетично значущих перетворень філогенезу - філембріогенези.Вони є відхиленнями від онтогенезу, характерного для предків, що виявляються в ембріогенезі, але мають адаптивне значення у дорослих форм. Так, закладки волосяного покриву з'являються у ссавців дуже ранніх стадіях ембріонального розвитку, але сам волосяний покрив має значення лише в дорослих організмів.
Такі зміни онтогенезу, будучи корисними, закріплюються природним відбором та відтворюються у наступних поколіннях. В основі цих змін лежать ті ж механізми, що зумовлюють вроджені вади розвитку: порушення проліферації клітин, їх переміщення, адгезії, загибелі чи диференціювання. Однак від вад їх так само, як і ценогенези, відрізняє адаптивна цінність, тобто. корисність та закріпленість природним відбором у філогенезі.
Залежно від того, на яких етапах ембріогенезу та морфогенезу конкретних структур виникають зміни розвитку, що мають значення філембріогенезів, розрізняють три їх типи.
1. Анаболії,або надставки, виникають після того, як орган практично завершив свій розвиток, і виражаються у додаванні додаткових стадій, що змінюють кінцевий результат.
До анаболій відносять такі явища, як придбання специфічної форми тіла камбалою лише після того, як з ікринки вилуплюється мальок, невідмінний від інших риб, а також поява вигинів хребта, зрощення швів у мозковому черепі, остаточний перерозподіл кровоносних судин та організму ссавця.
2. Девіації -ухилення, що виникають у процесі морфогенезу органу. Прикладом може бути розвиток серця в онтогенезі ссавців, у яких воно рекапітулює стадію трубки, двокамерну та трикамерну будову, але стадія формування неповної перегородки, характерної для плазунів, витісняється розвитком перегородки, побудованої і розташованої інакше і характерною тільки для ссавців. У розвитку легенів у ссавців також можна знайти рекапітуляцію ранніх стадій предків, пізніше морфогенез йде по-новому.
3. Архаллаксиси -зміни, які виявляються лише на рівні зачатків і виражаються у порушенні їх розчленування, ранніх диференціювань чи появі принципово нових закладок. Класичним прикладом архалаксису є
розвиток волосся у ссавців, закладка яких настає дуже ранніх стадіях розвитку і від початку відрізняється від закладок інших придатків шкіри хребетних.
За типом архаллаксису виникають хорда у примітивних безчерепних, хрящовий хребет у хрящових риб, розвиваються нефрони вторинної нирки у плазунів.
Зрозуміло, що з еволюції з допомогою анаболії в онтогенезах нащадків повністю реалізується основний біогенетичний закон, тобто. відбуваються рекапітуляції всіх предкових стадій розвитку. При девіаціях ранні предкові стадії рекапітулують, а пізніші замінюються розвитком у новому напрямку. Архаллаксиси повністю не допускають рекапітуляції у розвитку цих структур, змінюючи самі їх зачатки.
Якщо зіставити схему філембріогенезів з таблицею К. Бера, що ілюструє закон зародкової подібності, стане зрозуміло, що Бер вже був дуже близький до відкриття філембріогенезів, але відсутність еволюційної ідеї на його міркуваннях не дозволило більш ніж 100 років випередити наукову думку.
В еволюції онтогенезу найчастіше зустрічаються анаболії як філембріогенези, які лише малою мірою змінюють цілісний процес розвитку. Девіації як порушення морфогенетичного процесу в ембріогенезі часто відкидаються природним відбором і тому зустрічаються значно рідше. Найбільш рідко в еволюції проявляються архалаксиси у зв'язку з тим, що вони змінюють весь перебіг ембріогенезу, і якщо такі зміни зачіпають зачатки життєво важливих органів або органів, що мають значення ембріональних організаційних центрів, то часто вони несумісні з життям.
В одній і тій філогенетичній групі еволюція в різних системах органів може відбуватися за рахунок різних філембріогенезів.
Так, в онтогенезі ссавців простежуються всі етапи розвитку осьового скелета в підтипі хребетних (анаболії), у розвитку серця рекапітулують лише ранні стадії (девіація), а у розвитку придатків шкіри рекапітуляції взагалі відсутні (архаллаксис). Знання типів філембріогенезів в еволюції систем органів хордових необхідно лікарю для прогнозування можливості виникнення у плодів та новонароджених вроджених вад розвитку атавістичної природи. Дійсно, якщо в системі органів, що еволюціонує шляхом анаболії та девіацій, можливі атавістичні вади розвитку за рахунок рекапітуляції предкових станів, то у разі архалаксису це виключається повністю.
Крім ціногенезів та філембріогенезів в еволюції онтогенезу можуть виявлятися ще й відхилення часу закладення органів. гетерохронії -та місця їх розвитку - гетеротопії.Як перші, так і другі призводять до зміни взаємовідповідності структур, що розвиваються, і проходять жорсткий контроль природного відбору. Зберігаються лише ті гетерохронії та гетеротопії, які виявляються корисними. Прикладами таких адаптивних гетерохроній є зрушення у часі закладок найбільш життєво важливих органів у групах, що еволюціонують за типом арогенезу. Так, у ссавців, і особливо в людини, диференціювання переднього мозку значно випереджає розвиток інших відділів.
Гетеротопії призводять до формування нових просторових та функціональних зв'язків між органами, забезпечуючи надалі їх спільну еволюцію. Так, серце, що знаходиться у риб під ковткою, забезпечує ефективне надходження крові в зяброві артерії для газообміну. Переміщаючись у загрудинну область у наземних хребетних, воно розвивається і функціонує вже в єдиному комплексі з новими органами дихання - легенями, виконуючи і тут насамперед функцію доставки крові до дихальної системи газообміну.
Гетерохронії та гетеротопії залежно від того, на яких стадіях ембріогенезу та морфогенезу органів вони виявляються, можуть бути розцінені як філембріогенези різних типів. Так, переміщення зачатків головного мозку, що призводить до його вигину, характерного для амніот, і що проявляється на початкових етапах його диференціювання, є архаллаксисом, а гетеротопія сім'яника у людини з черевної порожнини через пахвинний канал в мошонку, що спостерігається в кінці ембріогенезу після окон - Типова анаболія.
Іноді процеси гетеротопії, однакові за наслідками, можуть бути філембріогенезами різних типів. Наприклад, у різних класів хребетних часто зустрічається переміщення поясів кінцівок. У багатьох груп риб, що ведуть придонний спосіб життя, черевні плавці (задні кінцівки) розташовуються допереду від грудних, а у ссавців і людини плечовий пояс і передні кінцівки в дефінітивному стані знаходяться значно каудальні місця їхньої початкової закладки. У зв'язку з цим іннервація плечового пояса у них здійснюється нервами, пов'язаними не з грудними, а з шийними сегментами спинного мозку. У згаданих вище риб черевні плавці іннервуються нервами не задніх тулубових, а передніх сегментів, розташованих вперед від центрів іннервації грудних плавців. Це свідчить про гетеротопію закладки плавників вже на стадії ранніх зачатків, у той час як переміщення переднього пояса кінцівок у людини відбувається на пізніших етапах, коли іннервація їх вже повністю здійснена. Вочевидь, у першому випадку гетеротопія є архаллаксис, тоді як у другому - анаболію.
Ціногенези, філембріогенези, а також гетеротопії та гетерохронії, виявившись корисними, закріплюються в потомстві та відтворюються в наступних поколіннях доти, доки нові адаптивні зміни онтогенезу не витіснять їх, замінивши собою. Завдяки цьому онтогенез як коротко повторює еволюційний шлях, пройдений предками, а й прокладає нові напрями філогенезу у майбутньому.
Ціногенез(від грец. kainós - новий і...генез).пристосування організму, що виникає на стадії зародка (плоду) або личинки і не зберігається у дорослої особини. Приклади Ц. - плацента ссавців, що забезпечує у плода дихання, харчування та виділення; зовнішні зябра личинок земноводних; яйцевий зуб у птахів, який служить пташенятам для пробивання шкаралупи яйця; органи прикріплення у личинки асцидій, плавальний хвіст у личинки трематод - церкарія та ін. Термін «Ц.» введений у 1866 Е. Геккелем для позначення тих ознак, які, порушуючи прояви палінгенезу , т. е. повторень далеких етапів філогенезу у процесі зародкового розвитку особини, неможливо простежити під час онтогенезу сучасних форм послідовність етапів філогенезу їхніх предків, т. е. порушують Біогенетичний закон. Наприкінці 19 ст. Ц. стали називати будь-яку зміну властивого предкам ходу онтогенезу (німецькі вчені Е. Менерт, Ф. Кейбель та ін.). Сучасне розуміння терміна "Ц." сформувалося в результаті робіт А. Н. Северцова, який зберіг за цим поняттям лише значення провізорних пристроїв, або ембріо-адаптацій. також Філембріогенез.
Ціногенез(грец. kainos новий + genesis зародження, освіта) - поява у зародка чи личинки пристосувань до умов існування, не властивих дорослим стадіям, напр. освіту оболонок у зародків вищих тварин.
або ж так
Філембріогенез (від грец. phylon - рід, плем'я, embryon - зародок і genesis - походження), еволюційна зміна онтогенезу органів, тканин і клітин, пов'язана як з прогресивним розвитком, так і з редукцією. Вчення про філембріогенез розроблено російським біологом-еволюціоністом А.М. Северцевим. Модуси (методи) філембріогенезу різняться за часом виникнення у розвитку цих структур.
Якщо розвиток певного органу у нащадків продовжується після тієї стадії, на якій воно закінчувалося у предків, відбувається анаболія (від грец. anabole – підйом) – надставка кінцевої стадії розвитку. Прикладом може бути формування чотирикамерного серця у ссавців. У земноводних серце трикамерне: два передсердя та один шлуночок. У плазунів у шлуночку розвивається перегородка (перша анаболія), проте ця перегородка у більшості з них неповна - вона лише зменшує перемішування артеріальної та венозної крові. У крокодилів та ссавців розвиток перегородки продовжується до повного поділу правого та лівого шлуночків (друга анаболія). У дітей іноді як атавізм міжшлуночкова перегородка буває недорозвиненою, що веде до тяжкого захворювання, що потребує хірургічного втручання.
Продовження розвитку органу не потребує глибоких змін попередніх стадій його онтогенезу, тому анаболія – найпоширеніший спосіб філембріогенезу. Попередні анаболія стадії розвитку органів залишаються порівнянними з етапами філогенезу предків (тобто є рекапітуляціями) і можуть служити для його реконструкції (див. Біогенетичний закон). Якщо розвиток органу на проміжних стадіях ухиляється від шляху, яким йшов його онтогенез у предків, відбувається девіація (від позднелат. deviatio - відхилення). Наприклад, у риб і у плазунів луски виникають як потовщення епідермісу і підстилаючого його сполучно-тканинного шару шкіри - коріуму. Поступово товщаючи, ця закладка вигинається назовні. Потім у риб коріум окостеніє, кісткова луска, що формується, протикає епідерміс і висувається на поверхню тіла. У плазунів, навпаки, кістка не утворюється, але епідерміс ороговіє, утворюючи рогові луски ящірок і змій. У крокодилів коріум може окостеніти, утворюючи кісткову основу рогових лусок. Девіації призводять до глибшої, ніж анаболії, перебудови онтогенезу, тому вони трапляються рідше.
Найрідше виникають зміни первинних зачатків органів - архаллаксиси (від грец. arche - початок і allaxis - зміна). При девіації рекапітуляцію можна простежити від закладення органу на момент ухилення розвитку. При архалаксисі рекапітуляції немає. Прикладом може бути розвиток тіл хребців у земноводних. У викопних земноводних - стегоцефалів і в сучасних безхвостих земноводних тіла хребців формуються навколо хорди з кількох, зазвичай трьох з боку тіла, окремих закладок, які потім зливаються, утворюючи тіло хребця. У хвостатих земноводних ці закладки не з'являються. Окостеніння розростається згори і знизу, охоплюючи хорду, отже відразу утворюється кісткова трубка, яка, товщаючи, стає тілом хребця. Цей архалаксис є причиною дискутованого питання про походження хвостатих земноводних. Одні вчені вважають, що вони походять безпосередньо від кістеперих риб, незалежно від інших наземних хребетних. Інші - що хвостаті земноводні дуже рано дивергували від решти земноводних. Треті, нехтуючи розвитком хребців, доводять близьку спорідненість хвостатих та безхвостих земноводних.
Редукція органів, що втратили своє адаптивне значення, також відбувається шляхом філембріогенезу, головним чином, за допомогою негативної анаболії - випадання кінцевих стадій розвитку. При цьому орган або недорозвивається і стає рудиментом, або зазнає зворотного розвитку та повністю зникає. Прикладом рудименту може бути апендикс людини - недорозвинена сліпа кишка, прикладом повного зникнення - хвіст пуголовків жаб. Протягом усього життя у воді хвіст росте, на його кінці додаються нові хребці та м'язові сегменти. Під час метаморфозу, коли пуголовок перетворюється на жабу, хвіст розсмоктується, причому процес йде у зворотному порядку - від кінця до основи. Філембріогенез – основний спосіб адаптивної зміни будови організмів у ході філогенезу.
Квиток 97
Принципи (методи) філогенетичних перетворень органів та функцій. Відповідність структури та функції у живих системах. Поліфункціональність Кількісні та якісні зміни функцій біологічних структур.
.
Принципи філогенетичних перетворень
органомназивають історично сформовану спеціалізовану систему тканин, що характеризується відмежованістю, сталістю форми, локалізації, внутрішньої конструкції шляхів кровообігу та іннервації, розвитком в онтогенезі та специфічними функціями. Будова органів часто дуже складна. Більшість їх поліфункціонально, тобто. виконує одночасно кілька функцій. У той самий час у реалізації будь-якої складної функції можуть брати участь різні органи. системою(кровоносна, видільна та ін.). Якщо одну і ту ж функцію виконує група органів різного походження, її називають апаратом.Прикладом служить дихальний апарат, що складається як з органів дихання, так і з елементів скелета і м'язової системи, що забезпечують дихальні рухи.
У процесі онтогенезу відбувається розвиток, а й заміна одних органів іншими. Органи зрілого організму називають дефінітивними;органи, що розвиваються та функціонують тільки в зародковому чи личинковому розвитку, - провізорними.Прикладами провізорних органів є зябра личинок земноводних, первинна нирка та зародкові оболонки вищих хребетних тварин (амніот).
В історичному розвитку перетворення органів можуть мати прогресивнийабо регресивнийхарактер. У першому випадку органи збільшуються у розмірах і стають складнішими за своєю будовою, у другому – зменшуються у розмірах, а їхня будова спрощується.
Якщо в двох організмів, що знаходяться на різних рівнях організації, виявляються органи, які побудовані за єдиним планом, розташовані в однаковому місці і розвиваються подібним чином з однакових ембріональних зачатків, це свідчить про спорідненість даних організмів. Такі органи називають гомологічними.Гомологічні органи часто виконують ту саму функцію (наприклад, серце риби, земноводного, плазуна і ссавця), але в процесі еволюції функції можуть і змінюватися (наприклад, передніх кінцівок риб і земноводних, плазунів і птахів).
При проживання неспоріднених організмів у однакових середовищах вони можуть виникати подібні пристосування, які у виникненні аналогічнихорганів. Аналогічні органи виконують однакові функції, будова їх, місце розташування та розвитку різко різні. Прикладами таких органів є крила комах та птахів, кінцівки та щелепний апарат членистоногих та хребетних.
Будова органів суворо відповідає функціям, що виконуються ними. При цьому в історичних перетвореннях органів зміна функцій обов'язково супроводжується зміною морфологічних характеристик органу.
Основним принципом еволюції органічних структур є принцип диференціації.Диференціація є поділ однорідної структури на відокремлені частини, які з різного становища, зв'язків коїться з іншими органами та різних функцій набувають специфічне будова. Таким чином, ускладнення структури завжди пов'язане з ускладненням функцій та спеціалізацією окремих частин. Диференційована структура виконує кілька функцій, і її будова складно.
Прикладом філогенетичної диференціації може бути еволюція кровоносної системи у вигляді хордових. Так, у представників підтипу безчерепних вона побудована дуже просто: одне коло кровообігу, відсутність серця та капілярів у системі зябрових артерій.
У надкласі риб є двокамерне серце і зяброві капіляри. У земноводних вперше з'являється розподіл кровоносної системи на два кола кровообігу, а серце стає трикамерним. Максимальна диференціація характерна для кровоносної системи ссавців, серце яких чотирикамерне, а в судинах досягається повне роз'єднання венозного та артеріального кровотоків.
Окремі частини диференціюється, раніше однорідної структури, спеціалізуючись у виконанні однієї функції, стають функціонально дедалі більше залежними з інших частин цієї структури і зажадав від організму цілому. Таке функціональне підпорядкування окремих компонентів системи в цілісному організмі називають інтеграцією.
Чотирикамерне серце ссавців є прикладом високоінтегрованої структури: кожен відділ виконує лише свою спеціальну функцію, яка не має ніякого сенсу у відриві від функцій інших відділів. Тому серце має автономну систему функціональної регуляції у вигляді парасимпатичного атріовентрикулярного нервового вузла і при цьому строго підпорядковане нейрогуморальній системі регуляції організму в цілому.
Отже, одночасно з диференціацією спостерігається і підпорядкування елементів цілісної системи організму, тобто. процес інтергації.
У процесі еволюції закономірним є як виникненнянових структур, так і їх зникнення.В основі лежить принцип диференціації, що проявляється на тлі первинної поліфункціональності та здатності функцій змінюватися кількісно. Будь-яка структура у своїй виникає з урахуванням попередніх структур незалежно від цього, якому рівні організації живого здійснюється процес филогенеза. Так, відомо, що близько 1 млрд років тому вихідний білок глобін слідом за дуплікацією вихідного гена диференціювався на міо- і гемоглобін - білки, що входять до складу відповідно м'язових і кров'яних клітин і диференціювалися у зв'язку з цим за функціями. У філогенезі центральної нервової системи хордових можна бачити диференціювання і зміну функцій структур: головний мозок формується з переднього кінця нервової трубки. Так само нові біологічні види утворюються у вигляді ізольованих популяцій вихідних видів (див. § 11.6), а нові біогеоценози - за рахунок диференціювання передіснуючих (див. § 16.2).
У зв'язку з тим, що нижче будуть розглянуті філогенези конкретних систем органів, докладніше зупинимося на закономірностях виникнення та зникнення органів. прикладом виникненняорганів служить походження матки плацентарних ссавців від парних яйцеводів При подовженні ембріонального розвитку ссавців виникає необхідність тривалішої затримки зародка в організмі матері. Це може здійснюватися тільки в каудальних відділах яйцеводів, порожнина яких при цьому збільшується, а стінка диференціюється таким чином, що до неї прикріплюється плацента, що забезпечує взаємозв'язок організму матері та плода. У процесі природного відбору зберігалися і успішно розмножувалися насамперед ті ссавці, в організмах самок яких потомство розвивалося найдовше. У результаті з'явився новий орган - матка, який би зародку оптимальні умови внутрішньоутробного розвитку (див. разд. 14.5.3) і підвищує виживання відповідних видів.
У виникненні такого складнішого і спеціалізованого органу, як око, спостерігаються самі закономірності. В основі формування органу зору, як і всіх органів чуття, лежать клітини шкірного епітелію, серед яких диференціюються і рецепторні, зокрема світлочутливі. Об'єднання їх у групи призводить до виникнення примітивних відокремлених органів зору, що дозволяють тваринам лише оцінювати освітленість. Занурення такого світлочутливого органу під шкіру забезпечує збереження ніжних клітин, але при цьому зорова функція може здійснюватися тільки завдяки прозорості покривів. Чутливість до світла примітивного органу зору посилюється при потовщенні прозорих покривів та придбанні ними здібностей заломлювати світло та фокусувати його промені на чутливих клітинах ока. Складний орган вимагає допоміжного апарату - захисних структур, м'язів, що приводять його в рух і т.д. Збільшений рівень складності організації ока з необхідністю супроводжується ускладненням регуляції його функцій, як і виявляється у посиленні його інтеграції як цілісної системи.
Зникнення,або редукція, орган у філогенезі може бути пов'язана з трьома різними причинами і має різні механізми. По-перше, орган, який раніше виконував важливі функції, може виявитися в нових умовах шкідливим. Проти нього спрацьовує природний відбір і орган досить швидко може повністю зникнути. Прикладів такого прямого зникнення органів небагато. Так, багато комах малих океанічних островів безкрилі внаслідок постійної елімінації їх популяцій літаючих особин вітром. Найчастіше спостерігається зникнення органів завдяки субституції новими структурами, виконують колишні функції з більшою інтенсивністю. Так зникають, наприклад, у плазунів та ссавців переваги та первинні нирки, замінюючись функціонально вторинними нирками. Так само у риб і земноводних відбувається витіснення хорди хребтом.
Найчастіший шлях до зникнення органів через поступове ослаблення їх функцій. Такі ситуації виникають зазвичай за зміни умов існування. Орган, який майже не виконує функцій, виходить з-під контролю природного відбору і виявляє зазвичай підвищену мінливість. Зміни, що виникають, викликають порушення корелятивних зв'язків з іншими частинами організму. Завдяки цьому такий орган часто стає шкідливим, і проти нього починає діяти природний відбір.
У медичній практиці відомо, що рудиментарні органи і в людини характеризуються широкою мінливістю. Треті великі корінні зуби, або «зуби мудрості», наприклад, характеризуються не лише значною варіабельністю будови та розмірів, але й різними термінами прорізування, а також особливою схильністю до карієсу. Іноді вони взагалі не прорізуються, а нерідко, прорізавшись протягом найближчих років повністю руйнуються. Те саме стосується і червоподібного відростка сліпої кишки (апендикса), який в нормі може мати довжину від 2 до 20 см і бути розташованим по-різному (за очеревиною, на довгій брижі, за сліпою кишкою і т.д.). Крім того, запалення апендикса (апендицит) зустрічається значно частіше, ніж запальні процеси в інших відділах кишківника.
Процес редукції органу протилежний до його нормального морфогенезу. Насамперед випадають закладки таких частин органу, які у нормі формуються останніми. При недорозвиненні кінцівок у людини зазвичай насамперед недорозвиваються фаланги I і V пальців, які закладаються останніми. У китоподібних, зовсім позбавлених задніх кінцівок завдяки послабленню їх функцій у філогенезі, все ж таки залишаються закладки елементів тазового пояса, що формуються в процесі морфогенезу найбільш рано.
Дослідження генетичних основ редукції органів показали, що структурні гени, що регулюють морфогенез, не зникають, тоді як істотних змін піддаються гени, що регулюють час закладки рудиментарних органів, або гени, відповідальні за феномен індукційних взаємодій у зародку, що розвивається. Дійсно, при пересадках мезодермального матеріалу дна ротової порожнини зародка ящірки в ротову порожнину курка, що розвивається, можливе формування у останнього зубів типової будови, а пересадка шкірної мезодерми ящірки під епідерміс спини курчата призводить до формування в ньому типових.
Онтогенез - це індивідуальний розвиток, комплекс процесів розвитку окремої особливо від утворення зиготи і до смерті. Розвиток відбувається завдяки реалізації генетичної інформації отриманої від батьків. Істотний вплив на її реалізацію надають умови довкілля. Філогенез - це історичний розвиток виду, еволюційний розвиток організмів. Обидва процеси тісно пов'язані. Знаючи напрями та перетворення органів та їх систем у процесі історичного розвитку, можна зрозуміти та пояснити виникаючі в процесі ембріогенезу аномалії розвитку.
Зв'язок отногенезу і філогенезу позначилася на низці біологічних законів і закномерностей.В 1828 Карл Бер сформулював три закони:
1. Закон Зародкової подібності – зародок якоїсь вищої тварини не буває схожий на іншу тварину, але схожий на її ембріон
2. закон послідовного появи ознак – найбільш загальні ознаки притаманні даної великої групи тварин виявляються в їх зародків раніше ніж ознаки спеціальні
3. закон мебріональної дивергенції – кожен зародок цієї форми тварин не проходить через інші форми а поступово відокремлюється від них.
Пояснити ці закони можна так що на ранніх етапах ембріогенезу зародки тварин різних класів хребетних (наприклад риби птиці ссавці) схожі між собою. Згодом між ними з'являються відмінності в межах класів, а далі – в межах загонів (приклад: зародки свині та людини)
Закон зародкової подібності К. Бера
У 1828 р. Карл фон Бер сформулював закономірність, яку називають Законом Бера: " Чим раніше стадії індивідуального розвитку порівнюються, тим більше подібності вдається виявити " . Порівнюючи стадії розвитку зародків різних видів та класів хордових, К. Бер зробив такі висновки.
Ембріони тварин одного типу на ранніх стадіях розвитку подібні.
Вони послідовно переходять у своєму розвитку від більш загальних ознак типу до більш приватних. В останню чергу розвиваються ознаки, що вказують на приналежність ембріона до певного роду, виду і, нарешті, індивідуальні риси.
Ембріони різних представників одного типу поступово відокремлюються один від одного.
Розвиток еволюційної ідеї надалі дозволило пояснити подібність ранніх зародків їх історичним спорідненістю, а придбання ними дедалі більше приватних рис із поступовим відокремленням друг від друга - дійсним відокремленням відповідних класів, загонів, сімейств, пологів та видів у процесі еволюції.
Зародкова схожість пояснюється тепер дійсним спорідненістю організмів, які поступове розбіжність (ембріональна дивергенція) служить очевидним відображенням історичного розбіжності даних форм (філогенетичної дивергенції). Отже, з індивідуального розвитку можна простежити історію цього виду.
Ціногенези - зміни в онтогенезах що призводять до відхилення від шляху предкових форм, пристосування, що виникають у зародків, личинок, що адаптує їх до довкілля. У дорослих організмів ценогенези зберігаються, тобто. виявляючись лише з ранніх стадіях онтогенезу, не змінюють типу організації дорослого організму, але забезпечують вищий рівень виживання потомства. Наприклад, для хобітів, як амніотичних організмів до ценогенезів відносять зародкові оболонки, жовтковий мішок та алантоїс, а для хобітів, як плацентарних тварин - ще й плацента з пуповиною
Філембріогенези - це ембріональні новоутворення, які мають філогенетичне значення. Час їх появи і способи різні (приклад з луски акули розвиваються: 1- рогові щитки рептилії шляхом девіації 2 перо птиці - анаболія 3 - волосся ссавця -архаллаксис. Еволюція частіше шляхом анаболії тому спостерігається рекапітуляція. Час появи ефіл -повністю змінюється розвиток органу.Рекапітуляції немає) 2- середні стадії(девіація-відхилення у розвитку) 3 - кінцеві стадії (анаболія-надставка у розвитку органу)
Людина та біосфера. Ноосфера – вищий етап еволюції біосфери. Реакції організму зміну екологічної ситуації. приклади. Правило Лібіха-Тінемана. Бочка Лібіха. Принцип Ле-Шательє-Браун.
Біосфера та людина. Сучасна людина сформувалася близько 30-40 тис. років тому. З цього часу в еволюції біосфери став діяти новий антропогенний чинник. Перша створена людиною культура палеоліт (кам'яний вік) тривала приблизно 20-30 тис. років; вона збіглася з тривалим періодом заледеніння. Економічною основою життя людського суспільства було полювання на великих тварин: шляхетного та північного оленя, шерстистого носорога, віслюка, коня, мамонта, туру. На стоянках людини кам'яної доби знаходять численні кістки диких тварин свідчення успішного полювання. Інтенсивне винищення великих травоїдних тварин призвело до порівняно швидкого скорочення їх чисельності та зникнення багатьох видів. Якщо дрібні травоїдні могли поповнювати втрати від переслідування мисливцями завдяки високій народжуваності, то великі тварини через еволюційну історію були позбавлені цієї можливості. Додаткові труднощі для травоїдних виникли внаслідок зміни природних умов наприкінці палеоліту. 10-13 тис. років тому настало різке потепління, відступив льодовик, ліси поширилися у Європі, вимерли великі тварини. Це створило нові умови життя, зруйнувало економічну базу людського суспільства, що склалася. Закінчився період розвитку, характеризовавшийся лише використанням їжі, тобто. суто споживчим ставленням до довкілля. У наступну епоху неоліту - поряд з полюванням (на коня, дику вівцю, благородного оленя, кабана, зубра, і т.д.), риболовлею і збиранням (молюски, горіхи, ягоди, плоди) все більшого значення набуває процес виробництва їжі. Робляться перші спроби одомашнення тварин та розведення рослин, зароджується виробництво кераміки. Вже 9-10 тис. років тому були поселення, серед залишків яких виявляють пшеницю, ячмінь, сочевицю, кістки свійських тварин кіз, овець, свиней. У різних місцях Передньої та Середньої Азії, Кавказу, Південної Європи розвиваються зачатки землеробського та скотарського господарства. Широко використовується вогонь для знищення рослинності в умовах підсічного землеробства, і як засіб полювання. Починається освоєння мінеральних ресурсів, зароджується металургія. Зростання населення, якісний стрибок у розвитку науки і техніки за останні два століття, особливо в наші дні, призвели до того, що діяльність людини стала фактором планетарного масштабу, що спрямовує сили подальшої еволюції біосфери. В.І. Вернадський вважав, вплив наукової думки та людської праці зумовило перехід біосфери в новий стан ноосферу (сферу розуму).
НООСФЕРА – ВИЩА СТАДІЯ РОЗВИТКУ БІОСФЕРИ
Сфера взаємодії суспільства та природи, в межах якої розумна
діяльність постає головним, визначальним фактором розвитку біосфери та
людства називається ноосферою.
Вперше термін "ноосфера" у 1926 – 1927 роках. вжили французькі вчені Еге.
Лекруа (1870 – 1954) та П. Тейяр де Шарден (1881 – 1955) у значенні "новий
покрив", "мислячий пласт", який, зародившись наприкінці третинного періоду,
розгортається поза біосферою над світом рослин та тварин. У їх
представлення ноосфера - ідеальна, духовна ("мисляча") оболонка Землі,
що виникла з появою та розвитком людської свідомості. Заслуга наповнення
даного поняття матеріалістичним змістом належить академіку В. І.
Вернадського (1965, 1978).
У поданні В. І. Вернадського, людина - частина живої речовини,
підпорядкованого загальним законом організованості біосфери, поза якою воно
існувати неспроможна. Людина є частиною біосфери, стверджував видатний
вчений. Метою суспільного розвитку має бути збереження організованості
біосфери. Однак збереження її первинної організованості - "недоторканою
природи" – не несе в собі творчого початку у потужну геологічну силу. "І
перед ним, перед його думкою та працею стає питання про перебудову біосфери
на користь вільно мислячого людства як єдиного цілого. Це нове
стан біосфери, до якого ми, не помічаючи цього, наближаємося, і є
"Ноосфера". Ноосфера є якісно новий етап еволюції
біосфери, у якому створюються нові форми її організованості як нове
єдність, що виникає внаслідок взаємодії природи та суспільства. У ній
закони природи тісно переплітаються із соціально-економічними законами
розвитку суспільства, утворюючи високу матеріальну цілісність "олюдненої"
природи".
В. І. Вернадський, який передбачив настання епохи науково-технічної
революції у XX столітті, основною передумовою переходу біосфери до ноосфери
вважав наукову думку. Матеріальним її виразом у перетворюваній людиною
біосферою є праця. Єдність думки та праці не тільки створює нову
соціальну сутність людини, а й визначає перехід біосфери в
ноосферу. "Наука є максимальна сила створення ноосфери" – таке головне
становище В. І. Вернадського у вченні про біосферу, що закликає перетворювати,
а не руйнувати ойкумену.
Реакції організму зміну екологічної ситуації.
Оптимальний вплив на різні організми той самий фактор може надавати при різних значеннях. Крім того, живі організми поділяють на здатні існувати в широкому або вузькому діапазонах зміни будь-якого фактора середовища. До кожного екологічного чинника організми пристосовуються щодо незалежним шляхом. Організм може мати пристосованість до вузького діапазону одного фактора і широкого діапазону - іншого. Для організму має значення як амплітуда, а й швидкість коливань тієї чи іншої чинника.
Якщо вплив умов середовища не досягає граничних значень, живі організми реагують на нього певними діями або змінами свого стану, що зрештою веде до виживання виду. Подолання несприятливих впливів тваринами можливе двома способами:
1) шляхом їх уникнення,
2) шляхом набуття витривалості. В основі реакцій у відповідь рослин лежить вироблення пристосувальних змін їх будови і процесів життєдіяльності. Мінливість - одне з основних якостей живого різних рівнях його організації. Генетична мінливість – основа спадкової мінливості ознак.
Правило мінімуму Лібіха
Згідно з правилом мінімуму Лібіха, урожай (продукція), його величина і стійкість у часі, управляється змінною ресурсів, таких як простір, час, речовина, енергія та різноманітність, що знаходиться в мінімумі. В даний час це правило поширене на функціонування різних об'єктів і загалом свідчить, що стан функції визначається тим фактором, що має мінімальне значення. У трактуванні Ю. Одума (1986), на організмовому рівні в стаціонарному стані лімітує буде життєво важлива речовина, доступні кількості якої найбільш близькі до необхідного мінімуму. Е. А. Мічерліхом було сформульовано правило сукупної дії факторів, що уточнює правило мінімуму Лібіха. Для тварин лімітуючими (обмежуючими) факторами, як правило, є наявність достатньої кількості корму, придатних укриттів (притулків), кліматичні умови.
Закон зародкової схожості.Дослідження початку ХIXстоліття вперше стали звертати увагу на подібність ранніх стадій розвитку ембріонів вищих тварин зі ступенями ускладнення організації, що ведуть від низькоорганізованих форм до прогресивних. Порівнюючи стадії розвитку зародків різних видів та класів хордових, К. Бер зробив такі висновки:
1. Ембріони тварин одного типу на ранніх стадіях розвитку подібні.
2. Вони послідовно переходять у своєму розвитку від більш загальних ознак типу до більш приватних. В останню чергу розвиваються ознаки, що вказують на приналежність ембріона до певного роду, виду і нарешті індивідуальні риси.
3. Ембріони різних представників одного типу поступово відокремлюються один від одного.
К. Бер, будучи еволюціоністом, було пов'язувати відкриті їм закономірності індивідуального розвитку з процесом филогенеза. Тому зроблені ним узагальнення мали значення лише емпіричних правил.
Розвиток еволюційної ідеї у подальшому дозволило пояснити подібність ранніх зародків їх історичним спорідненістю, а придбання ними дедалі більше приватних рис із поступовим відокремленням друг від друга – дійсним відокремленням відповідних класів, загонів, сімейств, пологів, видів у процесі еволюції.
Незабаром після відкриттів закону зародкової подібності Ч. Дарвін показав, що цей закон свідчить про спільність походження та єдності початкових етапів еволюції в межах типу.
Онтогенез – повторення філогенезу.Порівнюючи онтогенез ракоподібних з морфологією їх вимерлих предків, Ф. Мюллер, зробив висновки про те, що ракоподібні, що нині живуть, у своєму розвитку повторюють шлях, пройдений їх предками. Перетворення онтогенезу в еволюції, на думку Мюллера, здійснюється завдяки його подовженню за рахунок додавання до нього додаткових стадій або надставок. На основі цих спостережень, а також вивчення розвитку хордових Е. Геккель (1866) сформулював основний біогенетичний закон, відповідно до якого онтогенез є коротким і швидким повторенням філогенезу.
Повторення структур, притаманних предків, в ембріогенезі нащадків названо рекапітуляціями. Рекапітулують не тільки морфологічні ознаки – хорда, закладення зябрових щілин та зябрових дуг у всіх хордових, а й особливості біохімічної організації та фізіології. Так було в еволюції хребетних відбувається поступова втрата ферментів, необхідні розпаду сечової кислоти – препарату метаболізму пуринів. У більшості безхребетних кінцевий продукт розпаду сечової кислоти – аміак, у земноводних та риб – сечовина, у багатьох плазунів – алантоїн, а у деяких ссавців сечова кислота взагалі не розщеплюється та виділяється із сечею. В ембріогенезі ссавців та людини відзначені біологічні та біохімічні рекапітуляції: виділення ранніми зародками аміаку, пізніше сечовини, потім алантоїну, а на останніх стадіях розвитку – сечової кислоти.
Однак у онтогенезі високоорганізованих організмів який завжди спостерігається суворе повторення стадій історичного поступу, як це випливає з біогенетичного закону. Так, зародок людини ніколи не повторює дорослих стадій риб, земноводних, плазунів, а подібний по ряду рис лише з їх зародками. Ранні стадії розвитку зберігають найбільшу консервативність, завдяки чому рекапітулують повніше, ніж пізні. Це пов'язано з тим, що одним з найбільш важливих механізмів інтеграції ранніх етапів ембріогенезу є ембріональна індукція, а структури зародка, що формуються насамперед, такі як хорда, нервова трубка, ковтка, кишка та соміти, є організаційними центрами зародка, від яких залежить весь перебіг розвитку.
Генетична основа рекапітуляцій полягає в єдності механізмів генетичного контролю розвитку, що зберігається на базі загальних генів регуляції онтогенезу, які дістаються родинним групам організмів від спільних предків.
Онтогенез як основа філогенезу. Вчення А.Н.Северцова про філембріогенези. Анаболії, девіації та архалаксиси. Гетерохронії та гетеротопії біологічних структур в еволюції онтогенезу.
Спираючись лише на основний біогенетичний закон, неможливо пояснити процес еволюції: нескінченне повторення пройденого саме собою не народжує нового. Так життя існує Землі, завдяки зміні поколінь конкретних організмів, еволюція її протікає завдяки змін, які у їх онтогенезах. Ці зміни зводяться до того, що конкретні онтогенези відхиляються від шляху, прокладеного предковими формами, і набуває нових рис.
До таких відхилень відносяться, наприклад ценогенези - пристосування, що виникають у зародків або личинок і адаптують їх до особливостей довкілля. У дорослих організмів ціногенези не зберігаються. Прикладами ценогенезів є рогові утворення у роті личинок безхвостих земноводних, що полегшують їм живлення рослинною їжею. У процесі метаморфозу у жабки вони зникають, і система травлення перебудовується для харчування комахами і хробаками. До ценогенезів у амніот відносять зародкові оболонки, жовтковий мішок та алантоїс, а у плацентарних ссавців та людини – ще й плаценту з пуповиною.
Ціногенези, виявляючись лише на ранніх стадіях онтогенезу, не змінюють типу організації дорослого організму, але забезпечують більш високу ймовірність виживання потомства. Вони можуть супроводжуватися при цьому зниженням плодючості та подовженням зародкового або личинкового періоду, завдяки чому організм у постембріональному та постличинковому періоді розвитку виявляється більш зрілим та активним. Виникнувши та виявившись корисними, ценогенези відтворюватимуться і в наступних поколіннях. Так, амніон, що з'явився вперше у предків плазунів у кам'яновугільному періоді палеозойської ери, відтворюється у всіх хребетних, що розвиваються на суші, як у яйцекладних – плазунів та птахів, так і у плацентарних ссавців.
Інший тип філогенетично значимих перетворень філогенезу – філембріогенези. Вони є відхиленнями від онтогенезу, характерного для предків, що з'являються в ембріогенезі, але мають адаптивне значення у дорослих форм. Так, закладки волосяного покриву з'являються у ссавців дуже ранніх стадіях ембріонального розвитку, але сам волосяний покрив має значення лише в дорослих організмів. (філембріогенез – комплекс спадково обумовлених адаптивних перетворень онтогенезу).
Такі зміни онтогенезу, будучи корисними, закріплюються природним відбором та відтворюються у наступних поколіннях. В основі цих змін лежать ті ж механізми, що зумовлюють вроджені вади розвитку: порушення проліферації клітин, їх переміщення, адгезії, диференціювання чи загибель. Однак від пороків їх також, як і від ценогенезів, відрізняє адаптивна цінність, корисність і закріпленість природним відбором у філогенезі.
Залежно від того, на яких етапах онтогенезу та морфогенезу конкретних структур виникають зміни розвитку, що мають значення філембріогенезів, розрізняють їх три типи.
1. Анаболії, або надставки, виникають після того, як орган практично завершив свій розвиток, і виражаються у додаванні додаткових стадій, що змінюють кінцевий результат.
До анаболій відносяться такі явища, як набуття специфічної форми тіла рибою камбалою лише після того, як з ікринки вилуплюється мальок, невідмінний від інших риб, а також поява вигинів хребта, зрощення швів мозкового черепа, остаточний перерозподіл кровоносних судин в організмі ссавців. у морського півня (Trigla) спочатку розвиваються як і в інших видів риб, а потім відбувається анаболія - передні три промені плавця розростаються як пальцеподібні придатки. Плодові тіла грибів мають надставку для кращого поширення спор. Нирки вищих хребетних (припухлина, первинна та вторинна).
При еволюції шляхом анаболії попередня, колишня предків кінцевої стадії розвитку виявляється палінгенетичною рекапітуляцією. Вона відбиває попередній етап еволюції, організацію предків. Палінгенез Е. Геккель назвав ознаки зародка, що рекапітулюють ознаки предків.
2. Девіації або ухилення розвитку проміжних стадій онтогенезу (зміна шляху).
Прикладом девіації може бути розвиток луски в акулових риб і рептилій. У акул, як і у рептилій, спочатку виникають потовщення епідермісу, під якими накопичуються сполучнотканинні клітини коріуму. Ці закладки ростуть до поверхні тіла, поступово набуваючи форми лусок. У акул сполучнотканинні клітини коріуму служать основою для розвитку окостеніння і формувань кісткових лусок, що прободають епідерміс. У рептилій, навпаки, луски формуються за рахунок ороговіння епідермальних складок. Ймовірно, бульби та цибулини рослин сформувалися шляхом девіації з первинної ембріональної нирки. Розвиток серця в онтогенезі ссавців, у яких воно рекапітулює стадію трубки, двокамерну, трикамерну будову, але для стадії формування неповної перегородки, характерної для плазунів, витісняється розвитком перегородки, побудованої і розташованої інакше і характерною тільки для ссавців. У розвитку легенів у ссавців також виявляється рекапітуляція ранніх стадій предків, пізніше морфогенез йде іншим шляхом.
3. Архаллаксиси, чи зміна первинних зачатків. У цьому змінюється закладка органу, і з початку йде по іншому шляху, на відміну предків.
Наприклад, розвиток хребта у змій порівняно з ящірками. Число хребців у ящірок коливається від 30 до 35, у великих змій воно сягає 500 і більше. Подовження тіла у змій відбувається за рахунок закладки додаткових сомітів, що включають сегмент мускулатури та хребців. На одній і тій же стадії розвитку (судячи в порівнянні голови) формуються у гекона (ящірка) 24 соміту, а у вужа 34. На наступній стадії у гекона утворюється 42 соміту, а у вужа 142. Додаткові хребці вужа в порівнянні з хребцями гекона гомологів не мають.
Шляхом архаллаксису йде розвиток променів плавників у деяких риб, числа зубів у зубастих китів, розвиток волосся у ссавців, гомологічного ембріонального закладення луски риб та рептилій.
Таким чином, шляхом архалаксису в еволюції можуть виникати нові органи. Архаллаксис можна спостерігати лише у еволюції частин організму, але з організму як цілого. Редукція органів також відбувається за допомогою філембріогенезу. Сєверцов виділяв два основні типи редукції органів: рудиментація та афанізія.
✓ Рудиментація – повільне зникнення органу, що втратив функцію та став непотрібним. Такий орган в онтогенезі не розвивається повною мірою та довго зберігається у філогенезі. Наприклад, у печерної амфібії протеї очі закладаються і починають розвиватися як у всіх амфібій. Молоді личинки мають цілком нормальні очі. Потім темп росту очей уповільнюється, припиняється розвиток м'язів очей. У дорослої тварини очі виявляються маленькими, нерухомими, рудиментарними, розташованими під епідермісом.
✓ Афанізія – орган, розвинений та функціонуючий нормально у предків, у нащадків виявляється шкідливим. В цьому випадку орган може закладатися в онтогенезі, але потім його закладення повністю резорбується. Так відбувається редукція хвоста у пуголовків. Протягом усього личинкового розвитку цей орган зростає та розвивається. Дорослим жабам, що пересуваються стрибками, хвіст шкідливий, тому в період метаморфозу він редукується і зникає повністю.
Еге. Геккелем було показано, що зміни в онтогенезі в процесі еволюції можуть виникати за допомогою гетерохроній – усунення часу закладки того чи іншого органу чи структури, та гетеротопій – топографічного усунення місця закладки структури. Прикладом адаптивних гетерохроній є зрушення у часі закладення життєво важливих органів у ссавців та людини. Вони диференціювання переднього мозку істотно випереджає розвиток інших його відділів. Як приклади гетеротопій можна вказати на зміну місця закладки легень та плавального міхура, які первинно виникли з виростів, що лежать з боків кишечника; у нащадків легені перемістилися на черевну, а плавальний міхур – на спинну сторону кишечника, приклад – гетеротопія яєчка (анаболія), переміщення серця у хордових у філогенезі.
Завдяки ценогенезам, філембріогенезам, гетеротопії та гетерохронії, онтогенез не лише коротко повторює еволюційний шлях, пройдений предками, а й прокладає нові напрямки філогенезу у майбутньому.
Дослідники початку ХІХ ст. вперше почали звертати увагу на подібність стадій розвитку ембріонів вищих тварин із ступенями ускладнення організації, що ведуть від низькоорганізованих форм до прогресивних. У 1828 р. Карл фон Бер сформулював закономірність, яку називають Законом Бера: " Чим раніше стадії індивідуального розвитку порівнюються, тим більше подібності вдається виявити " . Порівнюючи стадії розвитку зародків різних видів та класів хордових, К. Бер зробив такі висновки.
1. Ембріони тварин одного типу на ранніх стадіях розвитку подібні.
2. Вони послідовно переходять у своєму розвитку від більш загальних ознак типу до більш приватних. В останню чергу розвиваються ознаки, що вказують на приналежність ембріона до певного роду, виду і, нарешті, індивідуальні риси.
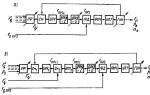
3. Ембріони різних представників одного типу поступово відокремлюються один від одного (рис. 1).
Малюнок 1. Подібність зародків різних класів хребетних на різних етапах
К. Бер, будучи еволюціоністом, було пов'язувати відкриті їм закономірності індивідуального розвитку з процесом филогенеза. Тому зроблені ним узагальнення мали значення лише емпіричних правил.
Розвиток еволюційної ідеї надалі дозволило пояснити подібність ранніх зародків їх історичним спорідненістю, а придбання ними дедалі більше приватних рис з поступовим відособленням друг від друга - дійсним відокремленням відповідних класів, загонів, сімейств, пологів та видів у процесі еволюції.
Еволюційне вчення, розроблене Ч. Дарвіном, яскраво висвітлило фундаментальне значення проблеми онтогенетичного розвитку. Зародкова схожість пояснюється тепер дійсним спорідненістю організмів, які поступове розбіжність (ембріональна дивергенція) служить очевидним відображенням історичного розбіжності даних форм (філогенетичної дивергенції). У зародку нащадків, писав Дарвін, бачимо «смутний портрет» предків. Отже, з індивідуального розвитку можна простежити історію цього виду.
Біогенетичний закон Е.Геккеля
Ф. Мюллер у роботі «За Дарвіна» (1864г.) сформулював думку, що зміни онтогенетичного розвитку, які у основі процесу еволюції, можуть виражатися у змінах ранніх чи пізніх стадій розвитку органів. У першому випадку зберігається лише загальна схожість молодих ембріонів. У другому випадку спостерігається продовження та ускладнення онтогенезу, пов'язане з надставкою стадій та повторенням (рекапітуляцією) в індивідуальному розвитку ознак більш далеких дорослих предків. Роботи Мюллера послужили основою для формулювання Е. Геккелем (1866) основного біогенетичного закону, згідно з яким онтогенез є коротким і швидким повторенням філогенезу. Тобто органічна особина повторює під час швидкого та короткого перебігу свого індивідуального розвитку найважливіші з тих змін форми, через які пройшли її предки під час повільного та тривалого перебігу їхнього палеонтологічного розвитку за законами спадковості та мінливості. Ознаки дорослих предків, що повторюються в ембріогенезі нащадків, він назвав палінгенезами. Пристосування до ембріональних чи личинкових стадій отримали назву ценогенезів.
Однак уявлення Геккеля сильно відрізнялися від поглядів Мюллера у питанні про співвідношення онтогенезу та філогенезу у процесі еволюції. Мюллер вважав, що нові форми виникають шляхом зміни ходу індивідуального розвитку, властивого їх предкам, тобто. зміни онтогенезу первинні стосовно філогенетичних змін. За Геккелем, навпаки, філогенетичні зміни передують змінам індивідуального розвитку. Еволюційно нові ознаки виникають над онтогенезу, а й у дорослого організму. Еволюціонує дорослий організм, і в процесі цієї еволюції ознаки зсуваються більш ранні стадії онтогенезу.
Таким чином, виникла проблема співвідношення онтогенезу та філогенезу, яка не вирішена й досі.
Геккель на відміну Мюллера вважав, що у філогенез впливають лише подовження онтогенезу шляхом надставки стадій, проте інші стадії залишаються без зміни. Отже, Геккель прийняв лише другий шлях історичних змін онтогенезу (за Мюллером) та залишив осторонь зміну самих стадій онтогенезу як основи філогенетичних перетворень. Саме на цій формі взаємозумовленості онтогенезу та філогенезу робили акцент Дарвін та Мюллер. Трактування біогенетичного закону у розумінні Мюллера пізніше була розвинена О.М. Северцова (1910-1939) в теорії філембріогенезів. Северцов поділяв погляди Мюллера про первинність онтогенетичних змін стосовно змін дорослих організмів і розглядав онтогенез як результат філогенезу, а й як його основу. Онтогенез як подовжується шляхом додавання стадій: він перебудовується у процесі еволюції; він має свою історію, закономірно пов'язану з історією дорослого організму та частково її визначальну.
Філембріогенези – це ембріональні зміни, пов'язані з філогенетичним розвитком дорослого організму. У процесі еволюції перебудовуються всі стадії розвитку. Нові зміни нерідко падають на останні стадії формоутворення. Онтогенез ускладнюється шляхом додавання або надставки стадій (анаболія). Тільки в цьому випадку є всі передумови для повторення в онтогенезі історичних етапів розвитку цих елементів у далеких предків (рекапітуляція). Онтогенез може, проте, змінюватися і будь-яких інших стадіях розвитку, відхиляючи у своїй всі пізні стадії від колишнього шляху (девіація). Зрештою, можлива й зміна самих зачатків органів або частин (архалаксис). Тоді весь онтогенез виявляється зміненим і в індивідуальному розвитку нащадків не зберігається вказівок на послідовність проходження історичних етапів розвитку їхніх предків.
Біологічна сутність біогенетичного закону Е. Геккеля
Біогенетичний закон Геккеля та теорія філембріогенезів Северцова відіграють важливу роль у розвитку морфології та самого еволюційного вчення. Вивчення індивідуального розвитку тварин дало достатньо доказів їхнього історичного розвитку. Біогенетичний закон є важливою складовою розробленого Е. Геккелем методу потрійного паралелізму, за допомогою якого проводять реконструкцію філогенезу. Цей метод заснований на зіставленні даних морфології, ембріології та палеонтології. Морфологи при реконструкції філогенезу досі користуються геккелівським принципом, згідно з яким онтогенез нащадків коротко повторює, рекапітулює етапи філогенезу предків. Спираючись лише на основний біогенетичний закон, неможливо пояснити процес еволюції: нескінченне повторення пройденого саме собою не народжує нового. Так як життя існує на Землі завдяки зміні поколінь конкретних організмів, еволюція її протікає завдяки змінам, що відбуваються в їх онтогенезі. Ці зміни зводяться до того, що конкретні онтогенези відхиляються від шляху, прокладеного предковими формами, і набувають нових рис.
До таких відхилень відносяться, наприклад, ценогенези - пристосування, що виникають у зародків або личинок і адаптують їх до особливостей довкілля. У дорослих організмів ціногенези не зберігаються. Прикладами ценогенезів є рогові утворення у роті личинок безхвостих земноводних, що полегшують їм живлення рослинною їжею. У процесі метаморфозу у жабки вони зникають і травна система перебудовується для живлення комахами та хробаками. До ценогенезів у плацентарних ссавців та людини - плаценту з пуповиною.
Ціногенези, виявляючись лише на ранніх стадіях онтогенезу, не змінюють типу організації дорослого організму, але забезпечують більш високу ймовірність виживання потомства. Вони можуть супроводжуватися при цьому зменшенням плодючості та подовженням зародкового або личинкового періоду, завдяки чому організм у постембріональному або післяличинковому періоді розвитку виявляється більш зрілим та активним. Виникнувши та виявившись корисними, ценогенези відтворюватимуться у наступних поколіннях.
Інший тип філогенетично значимих перетворень філогенезу – філембріогенези. Вони є відхиленнями від онтогенезу, характерного для предків, що виявляються в ембріогенезі, але мають адаптивне значення у дорослих форм. Так, закладки волосяного покриву з'являються у ссавців на дуже ранніх стадіях ембріонального розвитку, але сам волосяний покрив має значення тільки у дорослих організмів. В основі цих змін лежать ті ж механізми, які зумовлюють вроджені вади розвитку: порушення проліферації клітин, їх переміщення, адгезії, загибелі або диференціювання. корисність та закріпленість природним відбором у філогенезі.
Залежно від того, на яких етапах ембріогенезу та морфогенезу конкретних структур виникають зміни розвитку, що мають значення філембріогенезів, розрізняють три їх типи.
1. Анаболії, або надставки, виникають після того, як орган практично завершив свій розвиток, і виражаються у додаванні додаткових стадій, що змінюють кінцевий результат. До анаболій відносять такі явища, як придбання специфічної форми тіла камбалою лише після того, як з ікринки вилуплюється мальок, невідмінний від інших риб, а також поява вигинів хребта, зрощення швів у мозковому черепі, остаточний перерозподіл кровоносних судин та організму ссавця.
2. Девіації - ухилення, що виникають у процесі морфогенезу органу. Прикладом може бути розвиток серця в онтогенезі ссавців, у яких воно рекапітулює стадію трубки, двокамерну та трикамерну будову, але стадія формування неповної перегородки, характерної для плазунів, витісняється розвитком перегородки, побудованої і розташованої інакше і характерною тільки для ссавців. У розвитку легенів у ссавців також можна знайти рекапітуляцію ранніх стадій предків, пізніше морфогенез йде по-новому.
3. Архаллаксиси - зміни, що виявляються лише на рівні зачатків і виражаються у порушенні їх розчленування, ранніх диференціювань чи появі принципово нових закладок. Класичним прикладом архаллаксису є розвиток волосся у ссавців, закладка яких настає дуже ранніх стадіях розвитку і від початку відрізняється від закладок інших придатків шкіри хребетних. За типом архаллаксису виникають хорда у примітивних безчерепних, хрящовий хребет у хрящових риб, розвиваються нефрони вторинної нирки у плазунів.
Зрозуміло, що з еволюції з допомогою анаболії в онтогенезах нащадків повністю реалізується основний біогенетичний закон, тобто. відбуваються рекапітуляції всіх предкових стадій розвитку. При девіаціях ранні предкові стадії рекапітулують, а пізніші замінюються розвитком у новому напрямку. Архаллаксиси повністю не допускають рекапітуляції у розвитку цих структур, змінюючи самі їх зачатки.
В еволюції онтогенезу найчастіше зустрічаються анаболії як філембріогенези, які лише малою мірою змінюють цілісний процес розвитку. Девіації як порушення морфогенетичного процесу в ембріогенезі часто відкидаються природним відбором і тому зустрічаються значно рідше. Найбільш рідко в еволюції проявляються архалаксиси у зв'язку з тим, що вони змінюють весь перебіг ембріогенезу, і якщо такі зміни зачіпають зачатки життєво важливих органів або органів, що мають значення ембріональних організаційних центрів, то вони часто несумісні з життям.
Крім ценогенезів та філембріогенезів в еволюції онтогенезу можуть виявлятися ще й відхилення часу закладання органів – гетерохронії – та місця їх розвитку – гетеротопії. Як перші, так і другі призводять до зміни взаємовідповідності структур, що розвиваються, і проходять жорсткий контроль природного відбору. Зберігаються лише ті гетерохронії та гетеротопії, які виявляються корисними. Прикладами таких адаптивних гетерохроній є зрушення у часі закладок найбільш життєво важливих органів у групах, що еволюціонують за типом арогенезу. Так, у ссавців, і особливо в людини, диференціювання переднього мозку значно випереджає розвиток інших відділів.
Гетеротопії призводять до формування нових просторових та функціональних зв'язків між органами, забезпечуючи надалі їх спільну еволюцію. Так, серце, що знаходиться у риб під ковткою, забезпечує ефективне надходження крові в зяброві артерії для газообміну. Переміщаючись в загрудинну область у наземних хребетних, воно розвивається і функціонує вже в єдиному комплексі з новими органами дихання - легкими, виконуючи і тут насамперед функцію доставки крові до дихальної системи газообміну.
Гетерохронії та гетеротопії залежно від того, на яких стадіях ембріогенезу та морфогенезу органів вони виявляються, можуть бути розцінені як філембріогенези різних типів. Гетеротопія сім'яника у людини з черевної порожнини через паховий канал в мошонку, що спостерігається в кінці ембріогенезу після остаточного формування, - типова анаболія.
Ціногенези, філембріогенези, а також гетеротопії та гетерохронії, виявившись корисними, закріплюються в потомстві та відтворюються в наступних поколіннях доти, доки нові адаптивні зміни онтогенезу не витіснять їх, замінивши собою. Завдяки цьому онтогенез як коротко повторює еволюційний шлях, пройдений предками, а й прокладає нові напрями філогенезу у майбутньому .