突然変異の多様性と突然変異の種類。 遺伝と変動性
変動性- 生物が新しい特徴や性質を獲得する能力。 多様性のおかげで、生物は環境条件の変化に適応できます。
二つあります 変動の主な形態:遺伝性と非遺伝性。
遺伝的、 または 遺伝子型, 変動性- 遺伝子型の変化による生物の特性の変化。 さらに、それは組み合わせ性と突然変異性に分けられます。 組み合わせ変動は、配偶子形成および有性生殖中の遺伝物質 (遺伝子および染色体) の組み換えによって発生します。 突然変異の多様性は、遺伝物質の構造の変化の結果として発生します。
非遺伝的、 または 表現型、 または 修正, 変動性- 遺伝子型の変化によらない生物の特性の変化。
突然変異
突然変異- これらは、組織のさまざまなレベルでの遺伝物質の構造の永続的で突然の変化であり、生物の特定の特性の変化につながります。
「突然変異」という用語は、デ・フリーズによって科学に導入されました。 彼が作成した 突然変異理論、その主な規定は今日に至るまでその重要性を失っていない。
- 突然変異は、遷移を伴わずに突然、発作的に発生します。
- 突然変異は遺伝性です。 世代から世代へと永続的に受け継がれています。
- 突然変異は連続的な系列を形成せず、(修飾の変動性と同様に) 平均的なタイプに基づいてグループ化されず、質的な変化です。
- 突然変異には方向性がありません。どの遺伝子座でも突然変異が起こり、あらゆる方向にマイナーサインとバイタルサインの両方に変化を引き起こす可能性があります。
- 同じ変異が繰り返し発生する可能性があります。
- 突然変異は個別的なもの、つまり個々の個体で発生します。
突然変異が発生するプロセスは次のように呼ばれます。 突然変異誘発、そして突然変異を引き起こす環境要因は、 変異原性物質.
突然変異が発生した細胞の種類に応じて、それらは生成突然変異と体細胞突然変異に区別されます。
生成的突然変異生殖細胞で発生し、特定の生物の特性に影響を与えず、次の世代にのみ現れます。
体細胞突然変異体細胞で発生し、特定の生物体内で発現しますが、有性生殖中に子孫には伝達されません。 体細胞突然変異は、無性生殖(主に栄養生殖)によってのみ保存されます。
適応値に応じて、有益な変異、有害な変異(致死、半致死)、中立の変異に分類されます。 役に立つ- 活力を高め、 致死- 死を引き起こす 半致死的な- 活力を低下させ、 中性- 個人の生存能力には影響を与えません。 同じ突然変異でも、ある条件では有益な場合もあれば、別の条件では有害な場合もあることに注意してください。
発現の性質に応じて、突然変異が発生する可能性があります。 支配的なそして 劣性。 優性突然変異が有害である場合、個体発生の初期段階でその所有者の死を引き起こす可能性があります。 劣性突然変異はヘテロ接合体には現れないため、集団内に長期間「隠れた」状態で残り、遺伝的多様性の予備を形成します。 環境条件が変化すると、そのような突然変異を持った保因者が生存競争で有利になる可能性があります。
この突然変異を引き起こした突然変異原が特定されているかどうかに応じて、それらは区別されます。 誘発されたそして 自発的突然変異。 通常、自然突然変異は自然に発生しますが、誘導突然変異は人為的に引き起こされます。
突然変異が発生した遺伝物質のレベルに応じて、遺伝子、染色体、およびゲノムの突然変異が区別されます。
遺伝子変異
遺伝子変異- 遺伝子構造の変化。 遺伝子は DNA 分子の一部であるため、遺伝子変異はこの部分のヌクレオチド組成の変化を表します。 遺伝子変異は、以下の結果として発生する可能性があります。 1) 1 つ以上のヌクレオチドが他のヌクレオチドに置換される。 2) ヌクレオチドの挿入。 3)ヌクレオチドの損失。 4)ヌクレオチドの倍加。 5) ヌクレオチドの交互順序の変化。 これらの突然変異は、ポリペプチド鎖のアミノ酸組成の変化をもたらし、その結果、タンパク質分子の機能活性の変化をもたらします。 遺伝子変異により、同じ遺伝子の複数の対立遺伝子が生じます。
遺伝子の変異によって引き起こされる病気を遺伝病といいます(フェニルケトン尿症、鎌状赤血球貧血、血友病など)。 遺伝子疾患の遺伝はメンデルの法則に従います。
染色体の突然変異
これらは染色体の構造の変化です。 再配列は、1 つの染色体内で起こる可能性があります - 染色体内突然変異 (欠失、逆位、重複、挿入)、および染色体間で - 染色体間突然変異 (転座) です。
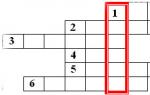
削除- 染色体セクションの喪失 (2); 反転— 染色体セクションの 180° 回転 (4, 5); 複製- 同じ染色体セクションの倍加 (3); 挿入— エリアの再配置 (6)。
染色体変異: 1 - パラ染色体。 2 - 削除。 3 - 重複。 4、5 - 反転。 6 - 挿入。
転座- 1 つの染色体の一部または染色体全体を別の染色体に移すこと。
染色体の突然変異によって引き起こされる病気は次のように分類されます。 染色体疾患。 このような疾患には、「猫の叫び声」症候群 (46、5p -)、ダウン症候群の転座変異型 (46、21 t21 21) などが含まれます。
ゲノム変異染色体の数の変化といいます。 ゲノム変異は、有糸分裂または減数分裂の正常な過程が破壊された結果として発生します。
一倍性- 染色体の完全な半数体セットの数の減少。
倍数性- 染色体の完全な一倍体セットの数の増加: 三倍体 (3 n)、四倍体 (4 n)など。
異倍性 (異数性) - 染色体の数の複数の増加または減少。 ほとんどの場合、染色体の数が 1 つずつ減少または増加します (まれに 2 つ以上になることもあります)。
異倍性の原因として最も可能性が高いのは、両親の一方の減数分裂中に相同染色体のペアが不分離であることです。 この場合、結果として生じる配偶子の一方には 1 つ少ない染色体が含まれ、もう一方には 1 つ多くの染色体が含まれます。 受精中にこのような配偶子と正常な一倍体配偶子が融合すると、特定の種に特徴的な二倍体セットと比較して、より少ないまたはより多くの染色体を持つ接合子が形成されます。 ヌロソミア (2n - 2), モノソミー (2n - 1), トリソミー (2n + 1), 四染色体 (2n+2)など
以下の遺伝図は、クラインフェルター症候群またはターナー・シェレシェフスキー症候群の子供の誕生が、母親または父親の減数分裂後期第 1 期における性染色体の不分離によって説明できることを示しています。
1) 母親の減数分裂中の性染色体の不分離
| R | ♀46,XX | × | ♂46,XY | ||
| 配偶子の種類 | 24、XX 24、0 | 23、X 23、Y | |||
| F | 47、XXX トリソミー X染色体上で |
47、XXY 症候群 クラインフェルター |
45、X0 ターナー症候群~ シェレシェフスキー |
45、Y0 死 受精卵 |
|
2) 父親の減数分裂中の性染色体の不分離
| R | ♀46,XX | × | ♂46,XY |
| 配偶子の種類 | 23、X | 24、XY 22、0 | |
| F | 47、XXY 症候群 クラインフェルター |
45、X0 ターナー症候群~ シェレシェフスキー |
ゲノム変異によって引き起こされる病気も染色体のカテゴリーに分類されます。 彼らの相続はメンデルの法則に従いません。 前述のクラインフェルター症候群またはターナー・シェレシェフスキー症候群に加えて、そのような疾患には、ダウン症候群 (47、+21)、エドワーズ症候群 (+18)、パトー症候群 (47、+15) が含まれます。
倍数性植物の特徴。 倍数体の生成は植物育種において広く使用されています。
遺伝的変動性の相同系列の法則 N.I. バビロバ
「遺伝的に近い種や属は、規則性を持った同様の一連の遺伝的変動を特徴とするため、ある種内の一連の形態を知ることで、他の種や属にも類似の形態が存在することを予測できます。 属と種が一般システム内で遺伝的に近い位置にあるほど、それらの一連の変動における類似性はより完全になります。 植物の科全体は一般に、その科を構成するすべての属と種を通過する特定の変動サイクルによって特徴付けられます。」
この法則は、小麦、ライ麦、大麦、オート麦、キビなどを含むポア科の例で説明できます。 したがって、カリオプシスの黒色はライ麦、小麦、大麦、トウモロコシおよび他の植物に見られ、カリオプシスの細長い形状は研究された科のすべての種に見られます。 遺伝的変動における相同系列の法則により、N.I. 自身がそれを可能にしました。 ヴァヴィロフは、小麦におけるこれらの特徴の存在に基づいて、これまで知られていなかったライ麦の多くの形態を発見しました。 これらには、芒のある穂と芒のない穂、赤、白、黒、紫色の穀物、粉っぽい穀物とガラス質の穀物などが含まれます。
| 形質の遺伝的変異 * | ライ麦 | 小麦 | オオムギ | オーツ麦 | アワ | ソルガム | トウモロコシ | 米 | ウィートグラス | ||
| トウモロコシ | 着色 | 黒 | + | + | + | — | — | + | + | + | + |
| 紫 | + | + | + | — | — | + | + | + | — | ||
| 形状 | ラウンド | + | + | + | + | + | + | + | + | + | |
| 拡張された | + | + | + | + | + | + | + | + | + | ||
| バイオル。 兆候 | ライフスタイル | 冬作物 | + | + | + | + | + | ||||
| 春 | + | + | + | + | + | + | + | + | |||
* 注記。 「+」記号は、指定された形質を持つ遺伝型の存在を意味します。
N.I.を開く ヴァビロフの法則は植物だけでなく動物にも当てはまります。 したがって、白皮症は、さまざまなグループの哺乳類だけでなく、鳥や他の動物でも発生します。 足の指が短いことは、人間、牛、羊、犬、鳥に見られ、鳥には羽がない、魚には鱗、哺乳類には羊毛などがあります。
遺伝的変動の相同系列の法則は、特定の種には見られないが、密接に関連した種に特徴的な形態の存在を予測できるため、選択にとって非常に重要です。 さらに、所望の形態は野生で見つけることも、人工的な突然変異誘発を通じて得ることもできる。
人為的突然変異
自然界では自然突然変異誘発は常に発生しますが、自然突然変異はかなりまれに発生します。たとえば、ショウジョウバエでは、白目の突然変異は配偶子 1:100,000 の頻度で形成されます。
身体への影響が突然変異の出現につながる要因は、と呼ばれます 変異原性物質。 突然変異原は通常 3 つのグループに分類されます。 物理的および化学的変異原は、人工的に突然変異を生成するために使用されます。
誘導突然変異誘発は、育種のための貴重な出発材料の作成を可能にし、また、突然変異誘発因子の作用からヒトを保護する手段を作成する方法を明らかにするため、非常に重要です。
修飾の変動性
修飾の変動性- これらは、遺伝子型の変化によって引き起こされるのではなく、環境要因の影響下で生じる生物の特性の変化です。 生息地は生物の特性の形成に大きな役割を果たします。 それぞれの生物は、生物の形態学的および生理学的特性を変化させる可能性がある要因の作用を経験しながら、特定の環境で発達し、生息します。 彼らの表現型。
環境要因の影響による特性の変化の例としては、矢尻の葉の形状の違いがあります。水に浸した葉はリボンのような形をしており、水面に浮かんでいる葉は丸い形をしており、水面に浮かんでいる葉は丸い形をしています。空気は矢印の形をしています。 紫外線の影響下で、人は(アルビノではない場合)皮膚にメラニンが蓄積して日焼けしますが、肌の色の濃さは人によって異なります。
修飾の変動性は、次の主な特性によって特徴付けられます。1) 非遺伝性。 2)変化の集団的性質(同じ条件下に置かれた同じ種の個体は同様の特性を獲得する)。 3) 環境要因の影響に対する変化の対応。 4) 遺伝子型に対する変動限界の依存性。
環境条件の影響で兆候が変化する可能性があるという事実にもかかわらず、この変動は無制限ではありません。 これは、遺伝子型が形質の変化が起こり得る特定の境界を決定するという事実によって説明されます。 形質の変動の程度、または修飾変動の限界は、と呼ばれます。 反応規範。 反応規範は、さまざまな環境要因の影響下で、特定の遺伝子型に基づいて形成される生物の表現型の全体として表現されます。 一般に、量的形質 (草丈、収量、葉の大きさ、牛の乳量、鶏の産卵量) は、質的形質 (毛色、乳脂肪含有量、花) よりも反応率が高く、大きく変動する可能性があります。構造、血液型)。 反応規範の知識は農業実践にとって非常に重要です。
植物、動物、人間の多くの特性の変更の変動性は一般法則に従います。 これらのパターンは、個人のグループにおける特性の発現の分析に基づいて特定されます ( n)。 研究対象の形質の発現の程度は、サンプル集団のメンバー間で異なります。 研究対象の特性のそれぞれの特定の値は次のように呼ばれます。 オプションそして文字で示されます v。 個々の変異体の発生頻度は文字で示されます。 p。 サンプル集団における形質の変動性を研究する場合、研究対象の形質の指標の昇順に個人を並べた変動シリーズが編集されます。
たとえば、小麦を 100 穂取った場合 ( n= 100)、穂の中の小穂の数を数えます ( v) と、指定された数の小穂を持つ穂の数を指定すると、バリエーション系列は次のようになります。
| 変異体 ( v) | 14 | 15 | 16 | 17 | 18 | 19 | 20 |
| 発生頻度 ( p) | 2 | 7 | 22 | 32 | 24 | 8 | 5 |

バリエーションシリーズをベースに構成されており、 変化曲線— 各オプションの出現頻度のグラフ表示。
特性の平均値はより一般的であり、それから大きく異なる変動はあまり一般的ではありません。 いわゆる 「正規分布」。 通常、グラフ上の曲線は対称です。
特性の平均値は、次の式を使用して計算されます。
どこ M— 特性の平均値; ∑( v
変異を突然変異といいます突然変異の発生によって引き起こされます。 突然変異- これらは、身体の特定の特性の変化につながる遺伝物質の遺伝的変化です。
突然変異理論の主な規定は、1901 年から 1903 年にかけて G. De Vries によって開発されました。 そして要約すると次のようになります。
突然変異は、形質の個別の変化として突然発生します。 突然変異はさまざまな形で現れ、有益な場合もあれば有害な場合もあります。
突然変異は指示されたものではありません(自然発生的)。つまり、染色体のどの部分でも突然変異が起こり、軽微な兆候とバイタルサインの両方に変化を引き起こす可能性があります。
ゲノムの変化の性質に基づいて、ゲノム変異、染色体変異、遺伝子変異のいくつかのタイプが区別されます。
ゲノム変異(異数性および倍数性) は、細胞ゲノム内の染色体の数の変化です。
染色体の突然変異、または染色体再構成は、染色体の構造の変化として表現されます。
不足、または反抗、- これは染色体の末端部分の喪失です。
削除- 染色体中央部分の欠損 (ABEFG);
重複- 染色体の特定の領域に局在する一連の遺伝子の二重または複数の繰り返し (ABCDECDEFG)。
反転- 染色体セクションの 180° 回転 (ABEDCFG);
転座- 同じ染色体のもう一方の端または別の非相同染色体 (ABFGCDE) へのセクションの移動。
で 欠陥、分割、重複染色体上の遺伝物質の量が変化します。 表現型の変化の程度は、対応する染色体領域の大きさと、重要な遺伝子が含まれているかどうかによって異なります。 染色体再構成の例は、ヒトを含む多くの生物で知られています。 重度の遺伝病である「猫の叫び声」症候群(病気の赤ちゃんが発する音の性質にちなんで名付けられた)は、5番目の染色体の欠損によるヘテロ接合性によって引き起こされます。 この症候群には精神遅滞が伴います。
重複ゲノムの進化において重要な役割を果たします。なぜなら、それらは新しい遺伝子の出現の材料として機能することができるためです。これは、以前は同一であった 2 つのセクションのそれぞれで異なる突然変異プロセスが発生する可能性があるためです。
で 逆位と転座遺伝物質の総量は変わりませんが、その位置が変わるだけです。
遺伝子または点突然変異- DNA 分子内のヌクレオチド配列の変化の結果。 この遺伝子のヌクレオチド配列の結果として生じる変化は、mRNA の構造の転写中に再現され、リボソーム上での翻訳の結果として形成されるポリペプチド鎖のアミノ酸配列の変化につながります。
遺伝子変異の影響は非常に多様です。 それらのほとんどは(劣性遺伝子であるため)表現型には現れませんが、特定の遺伝子のたった 1 塩基の変化が表現型に重大な影響を与えるケースは数多くあります。 一例は鎌状赤血球貧血です。この病気は、ヘモグロビンの合成を担う遺伝子の 1 つにおけるヌクレオチドの置換によって人間に引き起こされます。 これは、血液中でそのようなヘモグロビンを含む赤血球が変形し(円形から鎌形に)、すぐに破壊されるという事実につながります。 この場合、急性貧血が発症し、血液によって運ばれる酸素の量が減少します。
(遺伝子の突然変異は、紫外線、電離放射線、化学的突然変異原などの影響で発生します。地球の背景電離放射線は特に悪影響を及ぼします。自然の背景放射線がわずかに(1/3)増加しただけでも、たとえば、核兵器実験の結果、重度の遺伝性障害を持つ人々が世代ごとにさらに2,000万人発生する可能性があります。チェルノブイリ原子力発電所の事故のような出来事がどれほど危険であるかを想像するのは難しくありません。ウクライナ、ベラルーシ、ロシアの国民だけでなく、全人類に対しても同様の姿勢を示している。)
人類は膨大な数の疑問に直面していますが、その多くはまだ答えられていません。 そして、人に最も近いものは、その人の生理機能に関連しています。 外部および内部環境の影響下での生物の遺伝的特性の持続的な変化が突然変異です。 この要素は自然変動の源であるため、自然選択の重要な部分でもあります。
非常に多くの場合、ブリーダーは生物の突然変異に頼ることになります。 科学では、突然変異をゲノム、染色体、遺伝といういくつかのタイプに分類します。
遺伝によるものは最も一般的であり、私たちが最も頻繁に遭遇するものです。 これは一次構造を変更することであり、したがって mRNA から読み取られるアミノ酸も変更されます。 後者は、DNA 鎖の 1 つと相補的に配置されます (タンパク質生合成: 転写と翻訳)。
突然変異の名前には当初突然の変更がありました。 しかし、この現象に関する現代的な考え方は 20 世紀になって初めて発展しました。 「突然変異」という用語自体は、1901 年にオランダの植物学者であり遺伝学者であり、その知識と観察によってメンデルの法則を明らかにした科学者であるウーゴ・デ・フリースによって導入されました。 現代の突然変異の概念を定式化し、突然変異理論を発展させたのも彼ですが、同時期の 1899 年に同胞のセルゲイ・コルジンスキーによって定式化されました。
現代遺伝学における突然変異の問題
しかし、現代の科学者は理論の各点について説明を行っています。
結局のところ、何世代にもわたって蓄積される特別な変化があります。 また、元の製品のわずかな歪みからなる顔の突然変異があることも知られるようになりました。 新しい生物学的特性の再発に関する規定は、遺伝子変異にのみ適用されます。
それがどれほど有害であるか有益であるかを決定するのは、遺伝子型環境に大きく依存することを理解することが重要です。 多くの環境要因は、厳密に確立された遺伝子の自己複製プロセスである遺伝子の順序を乱す可能性があります。
自然選択の過程で、人類は有用な特性を獲得しただけでなく、病気に関連する最も好ましい特性も獲得しませんでした。 そして人間という種は、病理学的症状の蓄積を通じて自然から受け取ったものの代償を払っています。
遺伝子変異の原因

突然変異誘発因子。 ほとんどの突然変異は体に悪影響を及ぼし、自然選択によって規制されている形質を破壊します。 すべての生物は突然変異を起こしやすい性質がありますが、突然変異誘発因子の影響下ではその数が急激に増加します。 これらの要因には、電離、紫外線、高温、多くの化合物、ウイルスなどが含まれます。
抗変異原性因子、つまり遺伝装置を保護する因子には、遺伝暗号の縮重、遺伝情報を持たない不要な部分 (イントロン) の除去、および DNA 分子の二本鎖が含まれます。
突然変異の分類

1. 複製。 この場合、コピーは鎖内の 1 つのヌクレオチドから DNA 鎖の断片と遺伝子自体に起こります。
2. 削除。 この場合、遺伝物質の一部が失われます。
3. 反転。 この変更により、ある領域が 180 度回転します。
4. 挿入。 単一のヌクレオチドから DNA および遺伝子の一部への挿入が観察されます。
現代世界では、動物と人間の両方でさまざまな兆候の変化の発現にますます直面しています。 突然変異は、経験豊富な科学者を興奮させることがよくあります。
ヒトにおける遺伝子変異の例

1. 早老症。 早老症は、最もまれな遺伝的欠陥の 1 つであると考えられています。 この突然変異は、体の早期老化として現れます。 ほとんどの患者は13歳に達する前に亡くなりますが、20歳まで命を救える患者は少数です。 この病気は脳卒中や心臓病を引き起こすため、ほとんどの場合、死因は心臓発作または脳卒中です。
2. ユナー・タン症候群 (YUT)。 この症候群は、影響を受けた人が四つん這いで動くという点で特徴的です。 通常、SUT の人々は最も単純で原始的な言語を使用し、先天性脳不全に苦しんでいます。
3. 多毛症。 「狼男症候群」「エイブラムス症候群」とも呼ばれます。 この現象は中世以来追跡され、記録されています。 多毛症になりやすい人は、特に顔、耳、肩に、通常を超える多毛症が特徴です。
4. 重度の複合免疫不全症。 この病気にかかりやすい人は、生まれた時点で、平均的な人が持つ効果的な免疫システムをすでに奪われています。 1976年にこの病気を有名にしたデイビッド・ヴェッター氏は、免疫システムを強化する手術が失敗に終わり、13歳で亡くなった。
5. マルファン症候群。 この病気は非常に頻繁に発生し、手足の不均衡な発達と関節の過度の可動性を伴います。 あまり一般的ではありませんが、肋骨の癒合によって表される偏位があり、胸部が膨らんだり沈んだりします。 ボトム症候群になりやすい人によくある問題は、脊椎の湾曲です。
突然変異の変動性は、突然変異の発生によって引き起こされる変動性です。 突然変異とは、染色体の構造の変化によって引き起こされる形質、器官、または特性の遺伝的変化です。
突然変異の分類:
表現型別:
1. 形態学的 - 器官の成長と変化の性質が変化します。 形態学的変異には、表現型に目に見える変化をもたらす変異が含まれます。 たとえば、ホモ接合状態にあるショウジョウバエの白色遺伝子の劣性変異は白い目の色を引き起こしますが、野生型遺伝子の優性対立遺伝子は自然集団のハエに固有の赤い目の色を制御します。
2. 生理的 – 活力が増加(減少)します。 生理学的変異には、生物の重要な機能やその発達に影響を及ぼし、血液循環、呼吸、人間の精神活動、行動反応などのプロセスの破壊につながる変異が含まれます。 たとえば、血友病は血液凝固プロセスの違反に関連する遺伝性疾患です。
3. 生化学 – 体内の特定の化学物質の合成を阻害または変化させます。 生化学的変異は、酵素の完全な停止から通常は不活性な代謝経路の組み込みまで、酵素の活性の変化のすべてのケースを統合する広範なグループです。 一例としては、微生物における栄養要求性に対する多数の突然変異が挙げられる。微生物の担体は、野生型生物(原栄養生物)とは異なり、生命に必要な物質(アミノ酸、ビタミン、核酸前駆体など)を独立して合成することができない。 生化学的変異には、DNA 複製、DNA 損傷の修復、遺伝物質の転写および翻訳に関与する酵素の合成を妨害するさまざまな変異も含まれます。
遺伝子型別:
1. 遺伝的 - 対応するタンパク質分子の合成をコードする特定の遺伝子の領域における DNA 分子の構造の変化。 人間における遺伝子変異の結果、鎌状赤血球貧血、色覚異常、血友病などの病気が発生します。 遺伝子の突然変異により、新しい遺伝子対立遺伝子が生じ、進化の過程に影響を及ぼします。
2. 染色体 - 染色体の破損に伴う染色体の構造の変化(核が放射線または化学物質にさらされた場合)。
3. ゲノムとは、1 つ、複数、または完全な半数体の染色体のセットの追加または喪失を引き起こす突然変異です。 さまざまな種類のゲノム変異は、異数性および倍数性と呼ばれます。
ゲノム変異は染色体の数の変化に関連しています。 たとえば、植物では、染色体の数の複数の変化である倍数性の現象が非常に頻繁に検出されます。 倍数体生物では、細胞内の染色体 n の半数体のセットは、二倍体のように 2 つ (2n) ではなく、はるかに多くの回数 (3n、4n、5n、最大 12n) 繰り返されます。 倍数性は、有糸分裂または減数分裂の過程における破壊の結果です。紡錘体が破壊されると、複製された染色体は分離せず、未分割の細胞内に残ります。 その結果、染色体数2nの配偶子が出現します。 このような配偶子が正常な配偶子 (n) と合併すると、子孫は 3 組の染色体を持つことになります。 ゲノム変異が生殖細胞ではなく体細胞で発生すると、倍数体細胞のクローン(株)が体内に出現します。 多くの場合、これらの細胞の分裂速度は、正常な二倍体細胞 (2n) の分裂速度よりも速いです。 この場合、急速に分裂する倍数体細胞が悪性腫瘍を形成します。 それが除去または破壊されない場合、急速な分裂により、倍数体細胞が正常な細胞と置き換わります。 このようにして、多くの形態のがんが発生します。 有糸分裂紡錘体の破壊は、放射線または多くの化学物質(突然変異原)の作用によって引き起こされる可能性があります。
動物の染色体が 1 本または 2 本増加すると、発育異常や生物の死につながります。 例: ヒトのダウン症候群は 21 トリソミーであり、細胞内に合計 47 本の染色体があります。 突然変異は、放射線、X 線、紫外線、化学薬品、熱を使用して人工的に得ることができます。
相続の可能性については、
1. 生成 - 生殖細胞で発生し、遺伝します。
2. 体細胞 - 体細胞で発生し、遺伝しません。
細胞内での局在化により:
1. 核 - 細胞の遺伝物質で発生した突然変異 - 核、ヌクレオチド(原核生物の場合)。
2. 細胞質 - 突然変異は細胞質で生じ、細胞質 DNA を含む構造 (葉緑体、ミトコンドリア、プラスミド) の一部として現れます。
35. 自然発生的および誘発的突然変異プロセス。 突然変異と作用機序の概念。 コルピンスキーとH.デ・フリースの突然変異理論。
突然変異誘発とは、突然変異が発生するプロセスです。
自然発生的(自然) - 紫外線、放射線、化学的突然変異原など、生物の遺伝物質に対する突然変異誘発性の環境要因の影響により自然条件で発生する突然変異(人間に依存しない)。
誘発(人工) - 外部および内部環境の突然変異誘発因子の特別な影響(特に人間によって引き起こされる)の影響下での遺伝的変化の発生。
変異原とは、突然変異を引き起こす要因です。
1. 物理的(放射線、放射、温度)。
2. 化学物質(アルコール、フェノール);
3. 生物学的(ウイルス)。
(染色体内で) 突然変異を引き起こす一連の出来事は次のとおりです。 DNA損傷が発生します。 DNA 損傷が正しく修復されないと、突然変異が発生します。 重要でない DNA フラグメント (イントロン) で損傷が発生した場合、または重要なフラグメント (エクソン) で損傷が発生し、遺伝コードの縮重により違反が発生しなかった場合、突然変異が形成されますが、その生物学的影響は重要ではありません。または表示されない場合があります。
ゲノムレベルでの突然変異誘発は、一部の染色体の逆位、欠失、転座、倍数性、異数性、倍加、三重化(多重重複)などにも関連する可能性があります。
現在、点突然変異の形成の性質とメカニズムを説明するためにいくつかのアプローチが使用されています。 一般に受け入れられているポリメラーゼ モデルでは、塩基置換変異が形成される唯一の理由は、DNA ポリメラーゼにおける散発的なエラーであると考えられています。 現在、この考え方は一般的に受け入れられています。
ワトソンとクリックは、自然突然変異誘発の互変異性モデルを提案しました。 彼らは、DNA 分子が水分子と接触すると、DNA 塩基の互変異性状態が変化する可能性があるという事実によって、自然発生的な塩基置換突然変異の出現を説明しました。
突然変異理論は遺伝学の基礎の 1 つです。 それは、G. De Vries (1901-1903) の著作で G. メンデルの法則が発見された直後に始まりました。 さらに以前に、ロシアの植物学者S.I.は、遺伝的性質の突然の変化についての考えに達しました。 コルジンスキー (1899) の著書「異質生成と進化」。 人生のほとんどを植物の突然変異の多様性の問題の研究に捧げたコルジンスキー・デ・フリースの突然変異理論について話すのは当然です。 当初、突然変異理論は遺伝的変化の表現型の発現に完全に焦点を当てており、その発現のメカニズムには事実上まったく注意が払われていませんでした。 G. De Vries の定義によれば、突然変異とは、遺伝形質における発作的で断続的な変化の現象です。 これまで、数多くの試みにもかかわらず、G. De Vries によって与えられたものより優れた突然変異の簡潔な定義はありませんが、欠点がないわけではありません。 両者とも、突然変異によって自然選択を回避して新しい種が生み出される可能性があると誤って信じていた。
コルジンスキー - H. デ・フリースの突然変異理論の主な規定:
1. 突然変異は突然起こる
2. 新しいフォームは安定しています
3. 突然変異は質的な変化である
4. 有益なことも有害なこともある
5. 変異の検出は分析対象の個体数に依存する
6. 同じ変異が繰り返し発生する
突然変異性突然変異の発生によって引き起こされる変動と呼ばれます。 突然変異- これらは、身体の特定の特性の変化につながる遺伝物質の遺伝的変化です。
突然変異理論の主な規定は、1901 年から 1903 年にかけて G. De Vries によって開発されました。 そして要約すると次のようになります。
- 突然変異は、形質の個別の変化として突然発生します。
- 新しい形式は安定しています。
- 非遺伝的変化とは異なり、突然変異は連続的な系列を形成しません。 それらは質的な変化を表します。
- 突然変異はさまざまな形で現れ、有益な場合もあれば有害な場合もあります。
- 突然変異を検出する確率は研究対象の個体数に依存します。
- 同様の突然変異が繰り返し発生する可能性があります。
- 突然変異は無指向性(自然発生的)です。つまり、染色体のどの部分でも突然変異が起こり、軽微な兆候とバイタルサインの両方に変化を引き起こす可能性があります。
ゲノム変化の性質上突然変異には、ゲノム、染色体、遺伝子など、いくつかの種類があります。
ゲノム変異(異数性および倍数性)- これは細胞ゲノム内の染色体の数の変化です (詳細)。
染色体の突然変異、 または 染色体再構成、染色体の構造の変化として発現され、光学顕微鏡で特定して研究することができます。 さまざまなタイプの再構成が知られています (通常の染色体 - ABCDEFG)。
- 不足、 または 反抗、染色体の末端部分の喪失です。
- 削除- 染色体中央部分の欠損 (ABEFG);
- 重複- 染色体の特定の領域に局在する一連の遺伝子の二重または複数の繰り返し (ABCDECDEFG)。
- 反転- 染色体セクションの 180° 回転 (ABEDCFG);
- 転座- 同じ染色体のもう一方の端または別の非相同染色体 (ABFGCDE) へのセクションの移動。
欠損、分裂、重複が起こると、染色体上の遺伝物質の量が変化します。 表現型の変化の程度は、対応する染色体領域の大きさと、重要な遺伝子が含まれているかどうかによって異なります。 染色体再構成の例は、ヒトを含む多くの生物で知られています。 重度の遺伝病である「猫の叫び声」症候群(病気の赤ちゃんが発する音の性質にちなんで名付けられた)は、5番目の染色体の欠損によるヘテロ接合性によって引き起こされます。 この症候群には精神遅滞が伴います。 この症候群の子供は通常、早期に死亡します。
重複は、以前は同一だった 2 つのセクションのそれぞれで異なる突然変異プロセスが発生する可能性があるため、新しい遺伝子の出現の材料として機能する可能性があるため、ゲノムの進化において重要な役割を果たします。
逆位および転座の間、遺伝物質の総量は同じままで、その位置のみが変化します。 変異体と元の形態との交配は困難であり、それらの F1 雑種はほとんどの場合不稔であるため、このような変異は進化において重要な役割も果たします。 したがって、ここでは元の形式を相互に交差させることのみが可能です。 このような突然変異体が好ましい表現型を持っている場合、それらは新種の出現の初期形態となる可能性があります。 ヒトでは、これらの突然変異はすべて病理学的状態を引き起こします。
遺伝子または点突然変異- DNA 分子内のヌクレオチド配列の変化の結果。 この遺伝子のヌクレオチド配列の結果として生じる変化は、mRNA の構造の転写中に再現され、リボソーム上での翻訳の結果として形成されるポリペプチド鎖のアミノ酸配列の変化につながります。 遺伝子内のヌクレオチドの追加、欠失、または再配列を伴う、さまざまな種類の遺伝子変異があります。 これらは、重複、余分なヌクレオチドのペアの挿入、欠失(ヌクレオチドのペアの損失)、ヌクレオチドのペアの反転または置換(AT ↔ GC、AT ↔ CG または AT ↔ TA)です。
遺伝子変異の影響は非常に多様です。 それらのほとんどは(劣性遺伝子であるため)表現型には現れませんが、特定の遺伝子のたった 1 塩基の変化が表現型に重大な影響を与えるケースは数多くあります。 一例は鎌状赤血球貧血です。この病気は、ヘモグロビンの合成を担う遺伝子の 1 つにおけるヌクレオチドの置換によって人間に引き起こされます。 これは、血液中でそのようなヘモグロビンを含む赤血球が変形し(円形から鎌形に)、すぐに破壊されるという事実につながります。 この場合、急性貧血が発症し、血液によって運ばれる酸素の量が減少します。 貧血は体力の低下を引き起こし、変異対立遺伝子のホモ接合性を持つ人々の心臓や腎臓の問題や早期死亡につながる可能性があります。
遺伝子の突然変異は、紫外線、電離放射線、化学的突然変異原などの影響で発生します。 私たちの地球のバックグラウンド電離放射線は、特に悪影響を及ぼします。 たとえば核兵器実験の結果として、自然バックグラウンド放射線がわずかに増加しただけでも(1/3)、重度の遺伝性疾患を患う人々が各世代にさらに2,000万人発生する可能性があります。 チェルノブイリ原子力発電所の事故のような出来事が、ウクライナ、ベラルーシ、ロシアの国民だけでなく、全人類にどれほどの危険をもたらすかを想像するのは難しくありません。