胚類似性の法則を定式化したのは誰ですか。 個体系統関係
胚類似性の法則 動物発生学
一般発生学: 用語辞典 - スタヴロポリ. O.V. ディレコバ、T.I. ラピナ. 2010 .
他の辞書で「胚類似性の法則」が何であるかを確認してください。
動物発生学 ベアの法則、生殖類似性の法則 - この法則は 1828 年にカール エルネスト フォン ベアによって定式化されました。動物の大きなグループに特徴的な共通の基本的特徴は、動物の発生過程よりも早い時期に現れます。 一般発生学: 用語辞典
生物遺伝の法則- ヘッケル ミュラー (「ヘッケルの法則」、「ヘッケルのミュラーの法則」、「ダーウィン ミュラーのヘッケルの法則」、「基本的な生物遺伝法則」としても知られています): あらゆる生物は、その個別の発生 (個体発生) において既知の範囲で繰り返します。 ..ウィキペディア
生物遺伝の法則- (ギリシャの伝記 - 生命と起源 - 起源から) フリッツ ミュラーとエルンスト ヘッケルによって最初に定式化された立場は、生物が卵から発達した個体までの個々の発達の過程で通過する多くの形態である... ... 哲学事典
動物の比較解剖学- 比較形態学、異なる体系的なグループの動物を比較することによって、器官とそのシステムの構造と発達のパターンを研究する科学。 臓器の構造をその機能と関連付けて比較すると、理解することができます... ... ソビエト大百科事典
動物の比較解剖学- 比較形態学。異なる体系の動物を比較することによって、器官とそのシステムの構造と発達のパターンを研究する動物形態学のセクション。 グループ。 臓器の構造とその機能を比較すると、次のことが可能になります... ... 生物事典
進化の証拠- 始祖鳥の化石、「... ウィキペディア」の出版直後に発見
マクロ進化- 有機世界は、新しい属の種から、新しい科の属からなど、大きな体系的な単位を形成するプロセスです。大進化は微進化と同じ推進力に基づいています。 ... ... ウィキペディア
発生学- そして; そして。 [ギリシャ語から 胚とロゴス教義] 胚の形成と発達を研究する生物学の分野。 E. 動物。 比較例e. ◁ 発生学的、ああ、ああ。 この研究。 * * * 発生学 (胚と...学から)、... ... の科学 百科事典
進化論- (進化論および進化論とも) 地球の生物圏、その構成要素である生物地球圏、および個々の分類群や種の歴史的漸進的発展を肯定する生物学における考え方と概念の体系。... ... ウィキペディア
進化論の教育の歴史- 古代の哲学体系に起源を持ち、その考え方は宇宙論的な神話に根ざしています。 科学界が進化論を認識するきっかけとなったのは、チャールズ・ダーウィンの著書「自然手段による種の起源」の出版でした。 ... ウィキペディア
個体発生 – あらゆる段階で発生する遺伝情報の実装。
個体発生は遺伝的に制御されるプロセスです。 個体発生中に、遺伝子型が実現され、表現型が形成されます。
個体発生は生物の個々の発達であり、その発生の瞬間から生命の終わりまで生物が経験する一連の連続的な形態学的、生理学的、生化学的変化です。 O.には成長、つまり体重の増加、そのサイズ、分化が含まれます。 「お」という言葉。 E. Haeckel (1866) が生物遺伝法を策定したときに導入しました。
O. の歴史的実証の最初の試みは、I. f. によって行われました。 メッケル。 O. と系統発生の関係の問題は、C. ダーウィンによって提起され、F. ミュラー、E. Haeckel ら 遺伝の変化に関連するすべての新しい進化的特性は O. で生じますが、生物の生活条件へのより良い適応に寄与するものだけが自然選択の過程で保存され、次の世代に伝えられます。進化の中で固定されています。 酸素の法則、原因、要因に関する知識は、植物、動物、人間の発達に影響を与える手段を見つけるための科学的根拠として役立ち、これは動植物の飼育や医学にとって最も重要です。 。
系統発生は生物の歴史的発展です。 この用語はドイツ語で導入されました。 生理学研究の主な課題は、動物、植物、微生物の進化的変化を再構築し、これに基づいてそれらの起源と、研究対象の生物が属する分類群間の関連関係を確立することです。 この目的のために、E. ヘッケルは、形態学、発生学、古生物学という 3 つの科学からのデータを比較することによって、研究対象の体系的なグループの歴史的発展の過程を再構築できる「トリプル並列処理」の方法を開発しました。
胚類似性の法則
19世紀初頭の探検家。 彼らは初めて、高等動物の胚の発生段階と、低組織形態から進歩的な形態へと至る組織の複雑さ段階との類似性に注目し始めた。 K. Baer は、脊索動物のさまざまな種およびクラスの胚の発生段階を比較して、次の結論を出しました。
1. 同じ種類の動物の胚は、発生の初期段階では類似しています。
2. 彼らは、その発達において、タイプのより一般的な特徴から、より具体的な特徴へと一貫して移行します。 最後に、胚が特定の属、種、そして最終的には個々の形質に属していることを示す兆候が現れます。
3. 同じ種類の異なる代表の胚は、徐々に互いに分離されます。
K.ベアは進化論者ではなかったので、発見した個体の発達のパターンを系統発生のプロセスと結びつけることができませんでした。 したがって、彼の一般化は経験則以上の意味を持たなかった。
その後の進化論の考え方の発展により、初期胚の類似性をそれらの歴史的血縁関係によって説明することが可能になり、それらが徐々に互いに分離しながらより具体的な特徴を獲得していく、つまり対応する階級、目、科、科などの実際の分離が説明できるようになりました。進化の過程にある属と種。
胚類似性の法則の発見直後、チャールズ・ダーウィンは、この法則が型内の進化の初期段階の共通の起源と統一性を証明していることを示しました。
生物遺伝の法則ヘッケル・ミュラー: それぞれの生物は、その個別の発達 (個体発生) において、祖先またはその種 (系統発生) から受け継がれた形態をある程度繰り返します。
個体発生 - 系統発生の繰り返し
F. ミュラーは、甲殻類の個体発生とその絶滅した祖先の形態を比較し、現生の甲殻類は発生過程で祖先がたどった道を繰り返すと結論づけました。 F. ミュラーによれば、進化における個体発生の変容は、それに追加の段階や拡張を加えることによって長くなることによって行われます。 これらの観察と脊索動物の発生の研究に基づいて、E. Haeckel (1866) は基本的な生物遺伝法則を定式化しました。 個体発生は系統発生の短く急速な繰り返しです。
子孫の胚発生における祖先の特徴的な構造の繰り返しを、 要約。すべての脊索動物の脊索、鰓裂、鰓弓などの形態学的特徴だけでなく、生化学的組織や生理学的な特徴も再現します。 したがって、脊椎動物の進化において、プリン代謝産物である尿酸の分解に必要な酵素が徐々に失われます。 ほとんどの無脊椎動物では、尿酸の分解の最終生成物はアンモニアであり、両生類と魚類では尿素であり、多くの爬虫類ではアラントインであり、一部の哺乳類では、尿酸はまったく分解されず、尿中に排泄されます。 哺乳類とヒトの胚形成では、生化学的および生理学的再現が注目されます。初期の胚によるアンモニア、後期の尿素、次にアラントイン、そして発生の最終段階での尿酸の放出です。
しかし、高度に組織化された生物の個体発生においては、生物発生の法則からわかるように、歴史的発展段階の厳密な繰り返しが常に観察されるわけではありません。 したがって、ヒトの胚は魚類、両生類、爬虫類、哺乳類の成体段階を繰り返すことはなく、多くの特徴がそれらの胚とのみ似ています。 開発の初期段階は依然として最も保守的であるため、後の段階よりも完全に要約されています。 これは、胚発生の初期段階を統合するための最も重要なメカニズムの 1 つが胚誘導であり、脊索、神経管、咽頭、腸、体節など、最初に形成される胚の構造が、胚の組織的中心であり、そこから発生の全過程が決まります。
再現の遺伝的基盤は、発生の遺伝的制御機構の統一性にあり、この機構は、共通の祖先から関連する生物群に受け継がれる、個体発生の制御のための共通の遺伝子に基づいて保存されています。
要約(ラテン語の recapitulatio - 繰り返しから) - 進化の発達の初期段階に特徴的な特性の個々の発達における繰り返しを示すために生物学で使用される概念。
チケット96。
系統発生の基礎としての個体発生。 セノジェネシス。 個体発生の自律性。 フィ胚形成。 門胚形成に関するA.N. セベルツォフの教義。 それらの発生のメカニズム。 個体発生の進化における生物学的構造の異時性と異所性。
生物遺伝の基本法則だけに頼って、進化の過程を説明することは不可能です。過ぎ去ったものの終わりのない繰り返し自体が、何か新しいものを生み出すわけではありません。 地球上の生命は特定の生物の世代交代によって存在するため、その個体発生の変化によってその進化が起こります。 これらの変化は、特定の個体発生が祖先の形態によって定められた経路から逸脱し、新しい特徴を獲得するという事実に要約されます。
このような逸脱には、たとえば、 セノジェネシス -胚または幼虫の中で起こり、それらを環境の特徴に適応させる適応。 成体の生物では、新生は保存されません。 共生生殖の例としては、尾のない両生類の幼虫の口の中に角質が形成され、これにより幼虫は植物性食物を食べやすくなります。 カエルの変態の過程で、それらは消滅し、昆虫や虫を食べるために消化器系が再構築されます。 羊膜類の共生形成には、胚膜、卵黄嚢、尿膜が含まれ、胎盤哺乳類とヒトでは胎盤と臍帯も含まれます。
個体発生の初期段階にのみ現れるセノジェネシスは、成体生物の組織のタイプを変更しませんが、子孫の生存の可能性を高めます。 それらは、生殖能力の低下と胚または幼虫期間の延長を伴う可能性があり、これにより、胚後または幼虫後発育期の生物はより成熟して活動的になることが判明します。 セノジェネシスは発生し、有用であることが証明されたため、次の世代でも再現されるでしょう。 したがって、古生代の石炭紀の爬虫類の祖先に初めて現れた羊膜は、卵生の爬虫類と鳥類、および有胎盤哺乳類の両方で、陸上で発達するすべての脊椎動物で再現されています。
系統発生の系統学的に重要な変換のもう 1 つのタイプは次のとおりです。 門胚形成。それらは、祖先に特有の個体発生からの逸脱を表し、胚発生で明らかになりますが、成体の形態では適応的な重要性を持ちます。 したがって、毛芽は哺乳動物では胚発生の非常に初期段階で現れますが、毛自体が重要なのは成体の生物においてのみです。
個体発生におけるそのような変化は、有用であるため、自然選択によって固定され、次の世代で再生産されます。 これらの変化は、先天奇形を引き起こすのと同じメカニズム、つまり細胞の増殖、運動、接着、死または分化の障害に基づいています。 ただし、セノジェネシスと同様に、それらは適応価値によって悪徳とは区別されます。 系統発生における自然選択による有用性と固定化。
胚形成と特定の構造の形態形成の段階に応じて、系統形成の重要性を持つ発生変化が発生し、3つのタイプが区別されます。
1. 同化作用、または拡張は、器官の発達が実質的に完了した後に発生し、最終結果を変更する追加の段階の追加によって表現されます。
同化作用には、ヒラメが卵から孵化して稚魚になった後にのみ他の魚と見分けがつかない特定の体型を獲得する現象や、背骨の湾曲の出現、脳頭蓋骨の縫合糸の融合、最終的な変形などの現象が含まれます。哺乳類と人間の体内の血管の再分布。
2. 偏差 -器官の形態形成の過程で生じる逸脱。 例としては、哺乳類の個体発生における心臓の発達が挙げられます。この発達では、管段階、二室構造、三室構造が再現されていますが、爬虫類の特徴である不完全な中隔の形成段階は、心臓の発達段階に置き換えられています。隔壁の構造と位置が異なり、哺乳類にのみ特徴的です。 哺乳類の肺の発達では、祖先の初期段階の再現も見られ、その後の形態形成は新しい方法で進行します。
3. アーカラキシス -原基のレベルで検出され、その分裂、初期分化の違反、または根本的に新しい原基の出現として表現される変化。 アーカラキシスの典型的な例は次のとおりです。
哺乳類の毛の発達は、その毛根が発生の非常に初期の段階で起こり、最初から他の脊椎動物の皮膚付属器の毛根とは異なります。
弓頭蓋骨のタイプによると、脊索は原始的な頭蓋骨のない魚に現れ、軟骨脊椎は軟骨魚に現れ、二次腎臓のネフロンは爬虫類に発達します。
子孫の個体発生における同化作用による進化の過程で、基本的な生物遺伝の法則が完全に実現されることは明らかです。 祖先の発達段階すべてが再現されます。 逸脱の場合、初期の祖先段階が再現され、その後の段階は新しい方向への発展に置き換えられます。 Archallaxis は、これらの構造の発展における再現を完全に妨げ、その基本を変えてしまいます。
系統発生の図を、胚類似性の法則を示す K. ベアの表と比較すると、ベアがすでに系統発生の発見に非常に近づいていたことが明らかになりますが、彼の推論には進化論的な考え方が欠けていたため、彼が科学的思考よりも100年以上先を行くことを許してはならない。
個体発生の進化において、同化は系統発生として最も頻繁に起こりますが、これは発生プロセス全体をほんのわずかしか変化させません。 胚形成における形態形成プロセスの違反としての逸脱は、多くの場合、自然選択によって拒否されるため、あまり一般的ではありません。 アーカラキシスは、胚形成の過程全体を変化させるという事実により、進化において最もまれに発生します。そして、そのような変化が重要な器官や、重要な胚組織の中心である器官の初歩に影響を与える場合、それらは生命と両立しないことが判明することがよくあります。
同じ系統発生グループでも、系統胚形成の違いにより、異なる器官系での進化が発生する可能性があります。
したがって、哺乳類の個体発生では、脊椎動物のサブタイプにおける軸骨格の発達のすべての段階が追跡され(同化)、心臓の発達では初期段階のみが再現され(逸脱)、皮膚付属器の発達では、まったく要約がありません(アーカラキシス)。 医師が胎児や新生児における隔世遺伝の先天奇形の発生の可能性を予測するには、脊索動物の器官系の進化における系統胚形成の種類に関する知識が必要です。 実際、同化と逸脱を通じて進化する器官系において、先祖の状態の再現により隔世的発達障害が起こり得るとすれば、アーカラキシスの場合にはこれは完全に排除される。
新生と門胚形成に加えて、器官形成の時期の逸脱は個体発生の進化にも見られます。 異時性 -そしてその発展の場所 - ヘテロトピア。 1つ目と2つ目の両方とも、発達中の構造の相互対応に変化をもたらし、自然選択の厳密な制御を受けます。 有用であることが判明した異時性と異性体のみが保存されます。 このような適応異時性の例としては、形成の種類に応じて進化する集団における最も重要な器官の形成時期の変化が挙げられます。 したがって、哺乳類、特に人間では、前脳の分化が他の部分の発達を著しく促進します。
ヘテロトピーは、器官間の新しい空間的および機能的接続の形成につながり、それらのさらなる共同進化を確実にします。 したがって、魚の咽頭の下に位置する心臓は、ガス交換のために鰓動脈への効率的な血液の流れを保証します。 陸生脊椎動物の胸骨後領域に移動すると、新しい呼吸器官である肺との単一複合体で発達して機能し、ここでも主にガス交換のために呼吸器系に血液を送達する機能を実行します。
ヘテロクロニーとヘテロトピーは、それらが胚形成と器官形態形成のどの段階で現れるかに応じて、異なるタイプの系統形成とみなすことができます。 したがって、羊膜類に特徴的であり、その分化の初期段階で現れる、脳の原始的な部分の運動が弓膜類の屈曲につながり、腹腔から鼠径管を通って人間の精巣に至るまでの精巣の異所性である。陰嚢、最終的な形成後の胚形成の終わりに観察される - 典型的な同化。
場合によっては、異所性のプロセスは、結果が同一であっても、異なるタイプの系統形成である可能性があります。 たとえば、さまざまな種類の脊椎動物において、四肢帯の動きが非常に一般的です。 底棲生活を送る魚類の多くのグループでは、腹鰭(後肢)が胸鰭の前に位置し、哺乳類やヒトでは、最終的な状態では肩帯や前肢が胸鰭のかなり尾側に位置します。彼らの初期編成。 この点において、それらの肩帯の神経支配は、胸部ではなく脊髄の頸部に接続されている神経によって実行されます。 上述の魚では、腹鰭は後体幹神経ではなく、胸鰭の神経支配中心の前方に位置する前節によって神経支配されています。 これは、ひれの形成における異所視が原始初期の段階ですでに存在していることを示していますが、人間の四肢の前帯の運動は、神経支配がすでに完全に完了している後期の段階で発生します。 明らかに、最初のケースではヘテロトピアがアーカラキシスを表し、2 番目のケースでは同化が表されます。
セノジェネシス、系統発生、異所性および異時性は、有用であることが証明されており、子孫において固定され、個体発生における新たな適応変化がそれらを置き換えて置き換えるまで、次の世代で再生産されます。 このおかげで、個体発生は私たちの祖先がたどった進化の道を一時的に繰り返すだけでなく、将来の系統発生の新しい方向性も切り開きます。
セノジェネシス(ギリシャ語のkainós - 新しい、そして... 創世記 (... 創世記を参照)胚(胎児)または幼虫の段階で発生し、成体には持続しない生物の適応。 c. の例は、胎児に呼吸、栄養、排泄を提供する哺乳類の胎盤です。 両生類の幼生の外えら。 鳥の卵歯。雛が卵の殻を破るために使用します。 ホヤ幼虫の付着器官、吸虫幼虫の遊泳尾 - セルカリアなど。用語「C.」 1866 年に E. Haeckel によって導入され、パリンジェネシスの症状を妨害する特徴を指定します。 , つまり、個体の胚発生の過程で遠い系統発生の段階を繰り返すことは、現代の形態の個体発生中にその祖先の系統発生の一連の段階を追跡することを不可能にし、つまり、それらは生物遺伝の法則に違反します。 19世紀末。 祖先に特徴的な個体発生の過程におけるあらゆる変化は、色と呼ばれるようになりました(ドイツの科学者E.メーナート、F.カイベルなど)。 「C」という用語の現代的な理解。 A. N. セベルツォフの研究の結果として形成されました。セベルツォフは、この概念に対して暫定的な装置、または胚適応の意味のみを保持しました。 フィリムブリオジェネシスも参照。
セノジェネシス(ギリシャ語のkainos new +ジェネシスの起源、形成)-たとえば、成体段階の特徴ではない生活条件への適応の胚または幼虫の出現。 高等動物の胚における膜の形成。
とか、ぐらい
系統発生(ギリシャ語のファイロン - 属、部族、胚 - 胚、およびジェネシス - 起源に由来)、器官、組織、細胞の個体発生における進化的変化であり、進行性の発達と縮小の両方に関連しています。 系統発生学の学説は、ロシアの進化生物学者 A.N. によって開発されました。 セベルツォフ。 門胚形成の様式(方法)は、これらの構造の発達中の発生時期が異なります。
子孫における特定の器官の発達が、祖先で終了した段階の後に続く場合、アナボリアが発生します(ギリシャ語のアナボレから-上昇)-発達の最終段階の延長。 一例は、哺乳類における 4 室の心臓の形成です。 両生類の心臓は 3 つの部屋、つまり 2 つの心房と 1 つの心室を持っています。 爬虫類では、心室に中隔が発達しますが(第一同化)、ほとんどの爬虫類ではこの中隔は不完全であり、動脈血と静脈血の混合を減少させるだけです。 ワニや哺乳類では、右心室と左心室が完全に分離するまで中隔の発達が続きます(第二同化)。 小児では、隔世遺伝として心室中隔が未発達で、外科的介入が必要な重篤な病気につながることがあります。
器官の発達を延長するには、その個体発生の前段階に大きな変化を与える必要がないため、同化は系統発生の最も一般的な方法です。 同化に先立つ器官発達の段階は、祖先の系統発生の段階と同等のままであり(つまり、それらは要約である)、その再構築に役立ちます(生物遺伝の法則を参照)。 中間段階での器官の発達が、その祖先でその個体発生が起こった経路から逸脱すると、逸脱が発生します(後期ラテン語の逸脱 - 逸脱から)。 たとえば、魚や爬虫類では、鱗は表皮とその下にある皮膚の結合組織層である真皮の肥厚として現れます。 徐々に厚くなり、この原基は外側に曲がります。 次に、魚では真皮が骨化し、形成された骨の鱗が表皮を突き破って体の表面に移動します。 逆に、爬虫類では骨は形成されませんが、表皮が角質化し、トカゲやヘビの角質の鱗が形成されます。 ワニでは、真皮が骨化して、角質の鱗の骨の基礎を形成することがあります。 逸脱は同化よりも個体発生のより深い再構築につながるため、あまり一般的ではありません。
器官の主要な基礎の変化であるarchallaxis(ギリシャ語のarche-始まりとallaxis-changeから)は、最も頻繁には発生しません。 逸脱の場合、器官の起源から発達の逸脱の瞬間までを再現して追跡できます。 アーカラキシスでは、再現はありません。 一例は、両生類の椎体の発達です。 化石両生類である剣頭類と現代の無尾両生類では、椎体は、体の両側にいくつかの、通常は 3 つある別々の原基の弦の周りに形成され、その後、それらが結合して椎体を形成します。 有尾両生類では、これらの原基は現れません。 骨化は脊索を覆って上下に成長し、すぐに骨管が形成され、それが肥厚して椎体になります。 このアーカラキシスは、尾のある両生類の起源について今も議論されている問題の理由です。 一部の科学者は、他の陸生脊椎動物とは無関係に、それらは葉鰭のある魚の直接の子孫であると信じています。 尾のある両生類は他の両生類から非常に早く分岐したという説もある。 さらに、脊椎の発達を無視したものでは、尾のある両生類と尾のない両生類の密接な関係が証明されています。
適応的意義を失った器官の減少は、主に負の同化作用、つまり発生の最終段階の喪失を通じて、門胚形成を通じても起こります。 この場合、臓器は発育が不十分で原始的になるか、逆発育して完全に消失します。 原始的な例としては、ヒトの虫垂、つまり未発達の盲腸が挙げられ、完全に消失した例としては、カエルのオタマジャクシの尻尾が挙げられます。 水中での一生を通じて、尾は成長し、その末端に新しい椎骨と筋肉の部分が追加されます。 変態中、オタマジャクシがカエルに変わるとき、尾は溶解し、そのプロセスは逆の順序で、つまり端から根元まで起こります。 系統発生は、系統発生中に生物の構造を適応的に変化させる主な方法です。
チケット97
器官や機能の系統変化の原理(方法)。 生命システムにおける構造と機能の対応関係。 多機能性 生物学的構造の機能における量的および質的な変化。
.
系統変換の原理
権限歴史的に発達した組織の特殊なシステムと呼ばれ、境界、形状の恒常性、局在化、循環および神経支配経路の内部構造、個体発生および特定の機能の発達によって特徴付けられます。 臓器の構造は非常に複雑であることがよくあります。 それらのほとんどは多機能です。 複数の機能を同時に実行します。 同時に、さまざまな器官が複雑な機能の実行に参加することもあります。複雑な機能を実行するために団結する、起源が似ている器官のグループを呼びます。 システム(循環、排泄など)同じ機能が異なる起源の臓器のグループによって実行される場合、それはと呼ばれます。 装置。一例は呼吸器であり、これは呼吸器自体と、呼吸運動を提供する骨格および筋肉系の要素の両方で構成されます。
個体発生の過程では、発達が起こり、多くの場合、一部の器官が他の器官に置き換わります。 成熟した生物の器官は次のように呼ばれます。 決定的;胚または幼虫の発育中にのみ発達し機能する器官 - 仮。暫定臓器の例としては、両生類の幼生のえら、主要な腎臓、高等脊椎動物 (羊膜類) の胚膜などがあります。
歴史的発展において、臓器の変化は プログレッシブまたは 退行的なキャラクター。 前者の場合、臓器のサイズが大きくなり、構造がより複雑になりますが、後者の場合、臓器のサイズが小さくなり、構造が単純になります。
組織の異なるレベルに位置する 2 つの生物が、単一の計画に従って構築され、同じ場所に位置し、同じ胚の原始から同様の方法で発達する器官を持っている場合、これはこれらの生物の関係を示します。 このような器官はこう呼ばれます 相同な。相同な器官は多くの場合同じ機能を果たしますが(たとえば、魚類、両生類、爬虫類、哺乳類の心臓)、進化の過程で機能が変化することがあります(たとえば、魚類と両生類の前肢、爬虫類と鳥類)。
無関係な生物が同じ環境に住んでいる場合、それらは同様の適応を発達させる可能性があり、それが外観に現れます。 似ている臓器。 同様の器官は同じ機能を果たしますが、その構造、位置、発達は大きく異なります。 そのような器官の例は、昆虫および鳥の羽、節足動物および脊椎動物の手足および顎装置である。
臓器の構造は、臓器が実行する機能に厳密に対応しています。 また、臓器の歴史的変遷においては、機能の変化には必ず臓器の形態的特徴の変化が伴う。
有機構造の進化の基本原理は、 差別化。分化とは、均質な構造を別々の部分に分割し、異なる位置、他の器官との接続、およびさまざまな機能により、特定の構造を獲得することです。 このように、構造の複雑化は、常に機能の複雑化や各部品の特殊化を伴います。 分化した構造はいくつかの機能を実行し、その構造は複雑です。
系統発生的な分化の例としては、脊索動物門における循環系の進化が挙げられます。 したがって、頭蓋骨のないサブタイプの代表者では、血液循環が1つの円で、鰓動脈系に心臓と毛細血管が存在しないという非常に単純な構造になっています。
このスーパークラスの魚は、2つの部屋からなる心臓と鰓の毛細血管を持っています。 両生類では初めて、循環系が 2 つの循環円に分割され、心臓が 3 室になりました。 最大限の分化は哺乳類の循環系の特徴であり、その心臓は 4 つの部屋からなり、血管内では静脈血流と動脈血流の完全な分離が達成されます。
分化した、以前は均質な構造の個々の部分は、1 つの機能の実行に特化していましたが、この構造の他の部分や生物全体に機能的に依存するようになりました。 生物全体におけるシステムの個々の構成要素のこの機能的従属は、 統合。
哺乳類の 4 室の心臓は、高度に統合された構造の一例です。各部門は独自の特別な機能のみを実行し、他の部門の機能から切り離されては意味がありません。 したがって、心臓は副交感神経房室神経節の形で自律的な機能調節システムを備えており、同時に身体全体の神経液性調節システムに厳密に従属しています。
したがって、分化と同時に、身体の統合システムに対する部分の従属も観察されます。 統合プロセス。
進化の過程では当然のことですが、 出現新しい構造とその 失踪。それは、一次多官能性と量的に変化する機能の能力を背景にして現れる微分の原理に基づいています。 この場合の構造は、系統発生のプロセスが生物の組織のどのレベルで行われるかに関係なく、以前の構造に基づいて生じます。 したがって、約10億年前、元のグロビンタンパク質は、元の遺伝子の複製後、それぞれ筋肉と血液細胞の一部であるミオタンパク質とヘモグロビンタンパク質に分化し、したがって機能によって区別されたことが知られています。 脊索動物の中枢神経系の系統発生では、構造の分化と機能の変化も見ることができます。つまり、脳は神経管の前端から形成されます。 同様に、新しい生物学的種は、元の種の孤立した集団の形で形成され(§ 11.6 を参照)、また、既存の生物種の分化により、新しい生物地殻変動(§ 16.2 を参照)が形成されます。
特定の器官系の系統発生については以下で検討するため、器官の出現と消失のパターンについてさらに詳しく説明します。 例 出現器官は有胎盤哺乳類の子宮の起源であり、一対の卵管から生じます。 哺乳類の胚発生の長期化に伴い、母親の体内で胚をより長く保持する必要性が生じています。 これは、卵管の尾側部分でのみ行うことができ、その腔が拡大され、胎盤がそれに付着するように壁が分化し、母親と胎児の関係が確保されます。 自然選択の過程で、まずその体内で子孫が成長する哺乳類が保存され、繁殖に成功しました。 その結果、新しい器官である子宮が出現し、胚に子宮内発育に最適な条件を提供し(セクション14.5.3を参照)、対応する種の生存率を高めます。
目のようなより複雑で特殊な器官の出現においても、同じパターンが観察されます。 視覚器官の形成は、すべての感覚器官と同様に、皮膚上皮細胞に基づいており、その中で受容体細胞、特に光感受性細胞が分化します。 それらをグループにまとめると、原始的な別々の視覚器官が出現し、動物は照明を評価することだけが可能になります。 このような光に敏感な器官を皮膚の下に浸漬することで、繊細な細胞の安全性が確保されますが、視覚機能は外皮の透明性の出現によってのみ達成されます。 原始的な視覚器官の光に対する感受性は、透明なカバーが厚くなり、光を屈折させ、その光線を目の敏感な細胞に集中させる能力を獲得するにつれて増加します。 複雑な臓器には、保護構造、それを動かす筋肉などの補助装置が必要です。 目の組織の複雑さのレベルの増加は、必然的にその機能の調節の複雑化を伴い、それは統合システムとしての統合の強化として表現されます。
失踪系統発生における臓器の減少には 3 つの異なる理由があり、そのメカニズムも異なります。 第一に、以前は重要な機能を果たしていた臓器が、新たな状態では有害であることが判明する可能性があります。 自然選択はそれに逆らって働き、臓器は非常に早く完全に消滅する可能性があります。 このように臓器が直接消失する例はほとんどありません。 したがって、小さな海洋島の多くの昆虫は、風によってその個体群から飛翔個体が絶えず排除されるため、羽がありません。 臓器の消失は、より強力に同じ機能を実行する新しい構造に置き換えられるために観察されることが多くなります。 したがって、たとえば爬虫類や哺乳類では、前芽と一次腎臓が消失し、機能的に二次腎臓に置き換えられます。 同様に、魚類や両生類では、脊索が脊椎によって移動します。
臓器の消失に至る最も一般的な経路は、その機能が徐々に弱まっていくことです。 このような状況は通常、生活条件が変化したときに発生します。 ほとんど機能を果たさない器官は自然選択の制御から逃れ、通常は変動性の増大を示します。 結果として生じる変化は、身体の他の部分との相関関係の破壊を引き起こします。 このため、そのような器官はしばしば有害になり、自然選択がそれに対して作用し始めます。
人間の初歩的な臓器には大きなばらつきがあるという特徴があることは、医療現場では広く知られています。 たとえば、第 3 の大きな大臼歯、または「親知らず」は、構造とサイズが大きく異なるだけでなく、生える時期が異なること、さらに虫歯になりやすいという特徴があります。 まったく噴火しないこともありますが、噴火しても数年後には完全に破壊されてしまうこともよくあります。 同じことが盲腸の虫垂(虫垂)にも当てはまります。通常、長さは 2 ~ 20 cm で、位置は異なります(腹膜の後ろ、長い腸間膜上、盲腸の後ろなど)。 さらに、虫垂の炎症(虫垂炎)は、腸の他の部分の炎症過程よりもはるかに頻繁に発生します。
器官縮小のプロセスは、通常の形態形成とは逆です。 まず第一に、通常は最後に形成される臓器のそのような部分のしおりが落ちます。 人の四肢が未発達な場合、通常、最後に発達する第 1 指と第 5 指の指骨が最初に発達します。 系統発生における後肢の機能の弱化により後肢が完全に欠如しているクジラ目では、形態形成の過程の非常に初期に形成される骨盤帯の要素の初歩がまだ残っています。
器官減少の遺伝的基盤に関する研究では、形態形成を制御する構造遺伝子は消失しないが、初歩的な器官の形成時期を制御する遺伝子、または発生中の胚における誘導相互作用の現象に関与する遺伝子は重大な変化を受けることが示されている。 実際、トカゲの胚の口腔底の中胚葉材料を発育中のヒナの口腔に移植すると、ヒナは典型的な構造の歯を形成することが可能であり、皮膚中胚葉の移植は可能である。ヒナの背中の表皮の下にトカゲがいると、羽毛の代わりに典型的な角質の鱗が形成されます。
個体発生は個体の発生、特に受精卵の形成から死に至る個体の発生プロセスの複合体です。 発達は両親から受け取った遺伝情報の実装によって起こります。 環境条件はその実施に大きな影響を与えます。 系統発生は種の歴史的発展、つまり生物の進化的発展です。 両方のプロセスは密接に関連しています。 歴史的発生の過程における器官とそのシステムの方向と変化を知ることで、胚発生中に生じる発生異常を理解し、説明することが可能になります。
音形成と系統発生の関係は、多くの生物学的法則とパターンに反映されており、1828 年にカール ベアは 3 つの法則を定式化しました。
1. 胚類似性の法則 - 高等動物の胚は別の動物とは似ていないが、その胚には似ている
2. 文字の順次出現の法則 - 特定の大きなグループの動物に特徴的な、より一般的な文字は、より特殊な文字よりも早く胚の中で明らかにされます。
3. 胚分岐の法則 - 動物の特定の形態の各胚は、他の形態を通過せず、徐々にそれらから分離します。
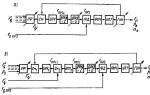
これらの法則は、胚発生の初期段階では、さまざまな種類の脊椎動物 (魚、鳥、哺乳類など) の胚が互いに似ているという方法で説明できます。 時間が経つにつれて、クラス内、さらには注文内でも差異が現れます (例: ブタとヒトの胚)。
K. ベアの生殖系列類似性の法則
1828 年、カール フォン ベアは、「個人の発達の初期段階を比較すると、より多くの類似点が見つかる」という、ベアの法則と呼ばれるパターンを定式化しました。 K. Baer は、脊索動物のさまざまな種およびクラスの胚の発生段階を比較して、次の結論を出しました。
同じ種類の動物の胚は、発生の初期段階では類似しています。
彼らは、その開発において、タイプのより一般的な特徴から、より具体的な特徴へと一貫して移行しています。 最後に、胚が特定の属、種、そして最終的には個々の形質に属していることを示す兆候が現れます。
同じ種類の異なる代表の胚は、徐々に互いに分離されます。
その後の進化論の考え方の発展により、初期胚の類似性をそれらの歴史的血縁関係によって説明することが可能になり、それらが徐々に互いに分離しながらより具体的な特徴を獲得していく、つまり対応する階級、目、科、科などの実際の分離が説明できるようになりました。進化の過程にある属と種。
現在、胚の類似性は生物間の実際の関係によって説明されており、それらの段階的な分岐(胚の分岐)は、これらの形態の歴史的な分岐(系統分岐)を明確に反映するものとして機能します。 したがって、特定の種の歴史は、個々の発達によって追跡することができます。
セノジェネシス - 祖先形態の経路からの逸脱につながる個体発生の変化、胚と幼虫で生じ、それらを環境に適応させる適応。 成体の生物では、新生は保存されません。 個体発生の初期段階でのみ発現するため、成体生物の組織の種類は変化しませんが、子孫の生存率を高めることができます。 たとえば、羊膜生物としてのホビットの場合、新生には胚膜、卵黄嚢、尿膜が含まれ、胎盤動物としてのホビットの場合、へその緒のある胎盤も含まれます。
系統発生は、系統発生学的に重要な胚性新生物です。 それらの出現の時期と方法は異なります(サメの鱗が発達する例: 1 - 偏向による爬虫類の角質皮膜 2 鳥の羽 - 同化 3 - 哺乳類の毛 - アーカラキシス。進化は同化によって起こることが多く、系統形成の出現時期: 1 形態形成の初期段階 (アーキラクシス - 器官の発達が完全に変化します。再現はありません。 2 - 中期段階 (逸脱 - 発達の逸脱) 3 - 最終段階 (アナボリア -臓器の発達をスーパーチャージします)
人間と生物圏。 ヌースフィアは、生物圏の進化の最高段階です。 環境状況の変化に対する体の反応。 例。 リービッヒ・ティーネマン則。 リービッヒバレル。 ル・シャトリエ・ブラウンの原則。
生物圏と人間。 現生人類は約3万年から4万年前に形成されました。 その時以来、生物圏の進化において新たな人為的要因が働き始めました。 人類が創造した最初の文化である旧石器時代(石器時代)は、約 2 万年から 3 万年続きました。 それは長い氷河期と一致していました。 人間社会の経済基盤は、アカシカやトナカイ、ケナガサイ、ロバ、馬、マンモス、オーロックスなどの大型動物の狩猟でした。 石器時代の人類遺跡では、狩猟が成功した証拠として野生動物の骨が多数発見されています。 大型草食動物の集中的な駆除により、その数は比較的急速に減少し、多くの種が絶滅しました。 小型草食動物が高い出生率のおかげで狩猟者による迫害による損失を補うことができたとしても、大型動物は進化の歴史によりその機会を奪われてきました。 旧石器時代末期の自然条件の変化により、草食動物にとってさらなる困難が生じました。 1万年から1万3千年前、急激な温暖化が起こり、氷河が後退し、森林がヨーロッパ全土に広がり、大型動物が絶滅した。 これにより新たな生活条件が生み出され、人間社会の既存の経済基盤が破壊されました。 食物の使用のみを特徴とするその発展の期間は終了しました。 環境に対する純粋に消費者の態度。 次の新石器時代には、狩猟(馬、野生の羊、アカシカ、イノシシ、バイソンなど)、釣り、採集(貝類、木の実、果実、果物)に加えて、食料生産のプロセスがますます重要になりました。 動物を飼いならし植物を飼育する最初の試みが行われ、陶器の生産が始まりました。 すでに9〜1万年前には集落があり、その遺跡からは小麦、大麦、レンズ豆、家畜の骨、ヤギ、羊、豚などが発見されています。 西アジア、中央アジア、コーカサス、南ヨーロッパのさまざまな場所で、農業と牛の繁殖の基礎が発展しています。 火は、焼畑農業における植生を破壊するためと狩猟の手段として広く使用されています。 鉱物資源の開発が始まり、冶金学が誕生します。 過去 2 世紀にわたる人口増加と科学技術の発展における質的飛躍は、特に今日では、人間の活動が地球規模の要素となり、生物圏のさらなる進化を導く力となっているという事実につながりました。 と。 ベルナツキーは、科学的思考と人間の労働の影響が生物圏の新しい状態、つまりヌースフィア(心の圏)への移行を決定すると信じていました。
ヌースフィア – バイオスフィア開発の最高段階
社会と自然との相互作用の範囲であり、その範囲内で合理的な
活動は生物圏の発展における主な決定要因であると思われます。
人類はヌースフィアと呼ばれています。
「ヌースフィア」という用語は 1926 年から 1927 年に初めて使用されました。 フランスの科学者 E.
ルクロワ (1870 – 1954) と P. テイヤール・ド・シャルダン (1881 – 1955) 「新しい」という意味
カバー」、「思考層」、これは第三紀の終わりに始まり、
生物圏の外側の動植物の世界に展開します。 彼らの中で
ヌースフィアは地球の理想的な精神的な(「思考」)殻です。
人間の意識の出現と発展とともに生まれました。 詰めるメリット
唯物論的な内容を含むこの概念は、アカデミアン V.I. に属します。
ヴェルナツキー (1965、1978)。
V.I. ヴェルナツキーの見解では、人間は生命体の一部であり、
生物圏の組織に関する一般法則に従い、その外では
存在できない。 人間は生物圏の一部であり、優れた生物圏であると主張
科学者。 社会発展の目標は組織を維持することであるべきです
生物圏。 ただし、その主要な組織は「無傷で」保存されています。
自然」 - それ自体の中に強力な地質学的力を生み出す創造原理を持ち込んでいません。
彼の前、彼の思考や仕事の前に、生物圏の再構築という問題が浮上します。
自由な思考を持つ人類全体の利益のために。 これは、新しい
私たちが気づかぬうちに近づいている生物圏の現状は、
「ヌースフィア」。 ヌースフィアは進化の質的に新しい段階を表している
生物圏、その組織の新しい形態が新しいものとして創造される
自然と社会の相互作用から生まれる一体性。 初期化
自然法則は社会経済法則と密接に絡み合っています
社会の発展、最高の物質的完全性を形成する「人間化された」
自然。"
科学と技術の時代の始まりを予言したV. I. ヴェルナツキー
20世紀の革命、生物圏からヌースフィアへの移行の主な前提条件
科学的思考と考えられます。 人間を改造したその物質的表現
生物圏は労働です。 思想と労働の団結は、新しいものを生み出すだけでなく、
人間の社会的本質だけでなく、生物圏の移行をあらかじめ決定するものでもあります。
ヌースフィア。 「科学はヌースフィアを創造する最大の力である」 - これが重要なことです
変革を求める生物圏の教義におけるV. I. ベルナツキーの立場、
エクメンを破壊しないでください。
環境状況の変化に対する体の反応。
同じ因子が、異なる値で異なる生物に最適な効果をもたらす可能性があります。 さらに、生物は、あらゆる環境要因の変化の広い範囲内で生存できるものと狭い範囲で存在できるものに分類されます。 生物は、比較的独立した方法で各環境要因に適応します。 生物は、ある要因の狭い範囲に適応する場合と、別の要因の広い範囲に適応する場合があります。 身体にとっては、特定の要素の振幅だけでなく振動速度も重要です。
環境条件の影響が極端な値に達しない場合、生物は特定の行動や状態の変化で環境条件に反応し、最終的には種の存続につながります。 動物による悪影響を克服するには、次の 2 つの方法があります。
1) それらを避けることで、
2)持久力を身につけることによって。 植物の反応の基礎は、その構造と生活プロセスにおける適応的な変化の発達です。 変動性は、組織のさまざまなレベルにおける生物の主要な特性の 1 つです。 遺伝的多様性は形質の遺伝的変動の基礎です。
リービッヒの最小規則
リービッヒの最小の法則によれば、収穫(生産)、その規模と長期にわたる安定性は、空間、時間、物質、エネルギー、多様性などの最小値である資源変数によって制御されます。 現在、この規則はさまざまなオブジェクトの機能に拡張されており、一般に関数の状態は最小値を持つ係数によって決定されると述べています。 Yu. Odum (1986) の解釈では、定常状態の生物レベルでは、制限物質は、利用可能な量が必要最小限に最も近い重要な物質となります。 E.A.ミッチャーリッヒは因子の結合作用の法則を定式化し、リービッヒの最小規則を明らかにしました。 動物の場合、原則として、十分な量の食料、適切な避難所(シェルター)、気候条件が制限要因となります。
胚類似性の法則。 19世紀初頭の研究では、高等動物の胚の発生の初期段階と、低組織形態から進歩的な形態に至る組織の複雑さの段階との類似性に初めて注目が集まり始めた。 さまざまな種および脊索動物のクラスの胚の発生段階を比較すると、 K. ベアは次の結論を出しました。
1. 同じ種類の動物の胚は、発生の初期段階では類似しています。
2. 彼らは、その発達において、タイプのより一般的な特徴から、より具体的な特徴へと一貫して移行します。 最後に、胚が特定の属、種、そして最終的には個々の形質に属していることを示す兆候が現れます。
3. 同じ種類の異なる代表の胚は、徐々に互いに分離されます。
K.ベアは進化論者ではなかったので、発見した個体の発達のパターンを系統発生のプロセスと結びつけることができませんでした。 したがって、彼の一般化は経験則以上の意味を持たなかった。
その後の進化論の考え方の発展により、初期胚の類似性をそれらの歴史的血縁関係によって説明することが可能になり、それらが徐々に互いに分離しながらより多くの特定の形質を獲得すること、つまり対応する階級、目、科、動物の実際の分離が説明できるようになりました。進化の過程にある属と種。
胚類似性の法則の発見直後、チャールズ・ダーウィンは、この法則が、ある型内での進化の初期段階の共通の起源と統一性を証明していることを示しました。
個体発生は系統発生の繰り返しです。 F. ミュラーは、甲殻類の個体発生とその絶滅した祖先の形態を比較し、現生の甲殻類は発生過程で祖先がたどった道を繰り返すと結論づけました。 ミュラーによれば、進化における個体発生の変容は、個体発生に追加の段階や拡張を加えることで長くなることによって行われる。 これらの観察と脊索動物の発生の研究に基づいて、E. Haeckel (1866) は基本的な生物遺伝法則を定式化しました。 個体発生 系統発生の簡潔かつ迅速な要約を表します。
子孫の胚形成における祖先に特徴的な構造の繰り返しを反復と呼びます。 すべての脊索動物の脊索、鰓裂および鰓弓の基部といった形態学的特徴だけでなく、生化学的組織および生理学的な特徴も再現します。 したがって、脊椎動物の進化において、プリン代謝産物である尿酸の分解に必要な酵素が徐々に失われます。 ほとんどの無脊椎動物では、尿酸の分解の最終生成物はアンモニアであり、両生類と魚類では尿素であり、多くの爬虫類ではアラントインであり、一部の哺乳類では、尿酸はまったく分解されず、尿中に排泄されます。 哺乳類とヒトの胚形成では、生物学的および生化学的な再現が注目されます。初期の胚によるアンモニア、後期の尿素、次にアラントイン、そして発生の最終段階での尿酸の放出です。
しかし、高度に組織化された生物の個体発生においては、生物発生の法則からわかるように、歴史的発展段階の厳密な繰り返しが常に観察されるわけではありません。 したがって、ヒトの胚は魚類、両生類、爬虫類の哺乳類の成体段階を繰り返すことはなく、多くの特徴がそれらの胚とのみ似ています。 開発の初期段階は依然として最も保守的であるため、後の段階よりも完全に要約されています。 これは、胚形成の初期段階を統合するための最も重要なメカニズムの 1 つが胚誘導であり、脊索、神経管、咽頭、腸、体節など、最初に形成される胚の構造が、それが依存する胚の組織的中心、発達の全過程。
再現の遺伝的基盤は、発生の遺伝的制御機構の統一性にあり、この機構は、共通の祖先から関連する生物群に受け継がれる、個体発生の制御のための共通の遺伝子に基づいて保存されています。
系統発生の基礎としての個体発生。 門胚形成に関するA.N. セベルツォフの教義。 アナボリア、逸脱、アーカラキシス。 個体発生の進化における生物学的構造の異時性と異所性。
生物遺伝の基本法則だけに頼って、進化の過程を説明することは不可能です。過ぎ去ったものの終わりのない繰り返し自体が、何か新しいものを生み出すわけではありません。 したがって、地球上に生命は特定の生物の世代交代によって存在し、個体発生の変化によってその進化が進行します。 これらの変化は、特定の個体発生が祖先の形態によって定められた経路から逸脱し、新しい特徴を獲得するという事実に要約されます。
このような逸脱には、例えば、共新生、つまり胚または幼虫の中で生じ、それらを環境の特性に適応させる適応が含まれます。 成体の生物では、新生は保存されません。 共生生殖の例としては、尾のない両生類の幼虫の口の中に角質が形成され、これにより幼虫は植物性食物を食べやすくなります。 カエルの変態の過程でカエルは消滅し、昆虫や虫を食べるために消化器系が再構築されます。 羊膜類の共生形成には、胚膜、卵黄嚢、尿膜が含まれ、胎盤哺乳類とヒトでは胎盤と臍帯も含まれます。
個体発生の初期段階にのみ現れるセノジェネシスは、成体生物の組織のタイプを変更しませんが、子孫の生存の可能性を高めます。 それらは、生殖能力の低下と胚または幼虫期間の延長を伴う可能性があり、これにより、胚後および幼虫後発育期の生物はより成熟して活動的になることが判明します。 発生し、有用であることが証明された後、セノジェネシスは次の世代でも再現されるでしょう。 したがって、古生代の石炭紀の爬虫類の祖先に初めて現れた羊膜は、卵生の爬虫類と鳥類、および有胎盤哺乳類の両方で、陸上で発達するすべての脊椎動物で再現されています。
系統発生学的に重要な系統発生のもう 1 つのタイプの変換は、系統発生です。 それらは祖先の個体発生の特徴からの逸脱を表し、胚発生に現れますが、成体の形態では適応的な重要性を持ちます。 したがって、毛芽は哺乳動物では胚発生の非常に初期段階で現れますが、毛自体が重要なのは成体の生物においてのみです。 (系統発生は個体発生の遺伝的に決定された適応的変換の複合体です)。
個体発生におけるそのような変化は、有用であるため、自然選択によって固定され、次の世代で再生産されます。 これらの変化は、先天奇形を引き起こすのと同じメカニズム、つまり細胞の増殖、移動、接着、分化、または死の障害に基づいています。 しかし、それらを悪徳や新生生成と区別するのは、それらの適応的価値、系統発生における自然選択による有用性と固定化です。
特定の構造の個体発生と形態形成の段階に応じて、系統発生の重要性を持つ発生変化が発生し、3つのタイプが区別されます。
1. 同化作用または拡張は、器官の発達が実質的に完了した後に発生し、最終結果を変える追加の段階の追加として表現されます。
同化作用には、ヒラメの魚が卵から孵化して稚魚になった後にのみ他の魚と区別がつかない特定の体型を獲得することや、背骨の湾曲の出現、脳頭蓋骨の縫合糸の融合などの現象が含まれます。哺乳類と人間の体内の血管の最終的な再配置、ホウボウ (トリグラ) の胸鰭は、他の魚種と同様に最初に発達し、その後同化が起こります。鰭の前の 3 つの条が指のような付属器のように成長します。 キノコの子実体は、胞子をより良く分布させるために延長されています。 高等脊椎動物の腎臓(前腎臓、一次および二次)。
同化を通じた進化では、以前の、以前の祖先の発達の最終段階は、古遺伝子の再現であることが判明します。 それは進化の前段階である祖先の組織を反映しています。 E. ヘッケルはパリンジェネシスを胎児の特徴、祖先の特徴を再現するものと呼びました。
2. 個体発生の中間段階の発達の逸脱、または回避(経路の変更)。
逸脱の例としては、サメ類や爬虫類の鱗の発達が挙げられます。 サメでは、爬虫類と同様に、表皮の肥厚が最初に現れ、その下に真皮の結合組織細胞が蓄積します。 これらの原石は体の表面に向かって成長し、徐々に鱗の形をとります。 サメでは、真皮の結合組織細胞は、骨化の発達と表皮に穴をあける骨鱗の形成の基礎として機能します。 逆に、爬虫類では、表皮のひだの角質化により鱗が形成されます。 植物の塊茎や球根は、一次胚芽からの逸脱によって形成された可能性があります。 哺乳類の個体発生における心臓の発達は、二室三室構造である管腔段階を再現していますが、爬虫類に特徴的な不完全な隔壁の形成段階では、心臓の発達によって置き換えられます。構造と位置が異なり、哺乳類にのみ特徴的な隔壁。 哺乳類の肺の発達では、祖先の初期段階の再現も見られ、その後の形態形成は異なる経路をたどります。
3. アーカラキシス、または一次原基の変化。 同時に、臓器の原基が変化し、その祖先とは異なり、最初から発達は異なる道をたどります。
たとえば、トカゲと比較したヘビの脊椎の発達。 トカゲの椎骨の数は 30 ~ 35 個ですが、大きなヘビでは 500 以上に達します。 ヘビの体の伸長は、筋肉や脊椎の一部を含む追加の体節の敷設によって起こります。 同じ発生段階(頭の比較から判断)で、ヤモリ(トカゲ)では24個の体節が形成され、草ヘビでは34個の体節が形成されますが、次の段階では、ヤモリでは42個、ヘビでは142個の体節が発生します。ヤモリの相同体の脊椎骨と比較して、グラスヘビの追加の脊椎骨はありません。
一部の魚類のひれ条の発生、ハクジラの歯の数、および魚類や爬虫類の鱗の胎児原基と相同な哺乳類の毛の発生は、アーカラシスによって起こります。
したがって、アーカラキシスを通じて、進化の過程で新しい器官が生じる可能性があります。 アーカラキシスは生物の一部の進化においてのみ観察され、生物全体としては観察されません。 器官の減少は、門胚形成によっても起こります。 セベルツォフ氏は、臓器縮小には主に 2 つのタイプがある、つまり初歩化と失語症であると特定しました。
✓ 初期化とは、機能を失い不要になった器官がゆっくりと消失することです。 このような器官は個体発生では完全には発達せず、系統発生では長期間存続します。 たとえば、洞窟の両生類プロテウスでは、他の両生類と同様に目が形成され、発達し始めます。 若い幼虫は完全に正常な目を持っています。 その後、目の成長速度が遅くなり、外眼筋の発達が止まります。 成体の動物の目は小さく、動かず、痕跡があり、表皮の下にあります。
✓ 失語症は、祖先では正常に発達し機能する器官ですが、子孫では有害であることが判明します。 この場合、器官は個体発生中に形成される可能性がありますが、その後、その原基は完全に吸収されます。 これはオタマジャクシの尾が縮小する方法です。 幼虫の成長を通じて、この器官は成長し、発達します。 ジャンプして移動する成体のカエルにとって、尾は有害であるため、変態中に尾は減少し、完全に消えます。
E. ヘッケルは、進化の過程における個体発生の変化が、ヘテロクロニー(特定の器官や構造の形成時期の変化)とヘテロトピア(構造が形成された場所の地形的変化)を通じて生じる可能性があることを示しました。 適応異時性の例としては、哺乳類やヒトにおける重要な器官の形成における時間の変化が挙げられます。 それらにおいては、前脳の分化が他の部分の発達を著しく促進します。 異所性の例として、肺と浮袋の位置の変化を挙げることができます。これは主に腸の側面にある増殖物から生じたものです。 子孫では、肺が腹側に移動し、浮き袋が腸の背側に移動しました。その例は、系統発生における脊索動物の心臓の動きである精巣異所視(アナボリア)です。
同生発生、系統発生、異所性および異時性のおかげで、個体発生は祖先がたどった進化の道を一時的に繰り返すだけでなく、将来の系統発生の新しい方向性を切り開きます。
19世紀初頭の探検家。 彼らは初めて、高等動物の胚の発生段階と、低組織形態から進歩的な形態へと至る組織の複雑さ段階との類似性に注目し始めた。 1828 年、カール フォン ベアは、「個人の発達の初期段階を比較すると、より多くの類似点が見つかる」という、ベアの法則と呼ばれるパターンを定式化しました。 K. Baer は、脊索動物のさまざまな種およびクラスの胚の発生段階を比較して、次の結論を出しました。
1. 同じ種類の動物の胚は、発生の初期段階では類似しています。
2. 彼らは、その発達において、タイプのより一般的な特徴から、より具体的な特徴へと一貫して移行します。 最後に、胚が特定の属、種、そして最終的には個々の形質に属していることを示す兆候が現れます。
3. 同じ種類の異なる代表の胚は、徐々に互いに分離されます(図 1)。
図 1. 異なる段階における異なるクラスの脊椎動物の胚の類似性
K.ベアは進化論者ではなかったので、発見した個体の発達のパターンを系統発生のプロセスと結びつけることができませんでした。 したがって、彼の一般化は経験則以上の意味を持たなかった。
その後の進化論の考え方の発展により、初期胚の類似性をそれらの歴史的血縁関係によって説明することが可能になり、それらが徐々に互いに分離しながらより多くの特定の形質を獲得すること、つまり対応する階級、目、科、動物の実際の分離が説明できるようになりました。進化の過程にある属と種。
チャールズ・ダーウィンによって開発された進化論は、個体発生の発生の問題の基本的な重要性を明確に強調しました。 現在、胚の類似性は生物間の実際の関係によって説明されており、それらの段階的な分岐(胚の分岐)は、これらの形態の歴史的な分岐(系統分岐)を明確に反映するものとして機能します。 ダーウィンは、私たちの子孫の胎児の中に、私たちの祖先の「漠然とした肖像」が見えると書いています。 したがって、特定の種の歴史は、個々の発達によって追跡することができます。
E. ヘッケルの生物遺伝法則
F. ミュラーは、著書「ダーウィンのために」(1864 年)の中で、進化の過程の根底にある個体発生の変化は、器官発達の初期または後期段階の変化として表現できるという考えを定式化しました。 最初のケースでは、若い胚の一般的な類似性のみが保存されます。 2番目のケースでは、より遠い成体祖先の特徴の個々の発達における段階の追加と反復(反復)に関連した個体発生の延長と複雑化が見られます。 ミュラーの研究は、E. ヘッケル (1866) による基本的な生物遺伝法則の定式化の基礎となり、それによれば、個体発生は系統発生の短くて急速な繰り返しであるとされています。 つまり、有機的個体は、遺伝と変動性の法則に従って、その祖先が古生物学的発達のゆっくりとした長い過程で経験した形態の最も重要な変化を、その個体の発達の急速かつ短期間の過程で繰り返す。 。 彼は、子孫の胚発生において繰り返される成人祖先の特徴をパリンジェネシスと呼びました。 胚または幼虫の段階への適応は、新生と呼ばれます。
しかし、ヘッケルの考えは、進化の過程における個体発生と系統発生の関係の問題に関するミュラーの見解とは大きく異なりました。 ミュラーは、進化的に新しい形態は、祖先の特徴である個々の発達の過程を変えることによって生じると信じていました。 個体発生の変化は系統発生の変化に関連して主要です。 ヘッケルによれば、逆に、系統発生の変化は個体の発達の変化に先行するという。 進化上の新しい形質は、個体発生の際ではなく、成体生物の中で生じます。 成体生物は進化しますが、この進化の過程で、特徴は個体発生の初期段階に移行します。
したがって、個体発生と系統発生の関係の問題が生じましたが、まだ解決されていません。
ヘッケルは、ミュラーとは異なり、系統発生は段階の追加による個体発生の拡張によってのみ影響を受け、他のすべての段階は変化しないと信じていました。 その結果、ヘッケルは(ミュラーによれば)個体発生における歴史的変化の第二の道のみを受け入れ、系統発生的変換の基礎としての個体発生の段階そのものの変化は脇に置いた。 ダーウィンとミュラーが強調したのは、個体発生と系統発生の間のこの形の相互依存性でした。 ミュラーによって理解された生物遺伝の法則の解釈は、後に A.N. によって発展されました。 セベルツォフ (1910-1939) の系統発生理論。 セベルツォフは、成体生物の変化に関連した個体発生的変化の優位性についてミュラーの見解を共有し、個体発生を系統発生の結果としてだけでなく、その基礎としても考慮した。 個体発生は段階を追加することで延長されるだけではなく、進化の過程で完全に再構築されます。 それには独自の歴史があり、成体の生物の歴史と自然に結びつき、それを部分的に決定します。
系統発生は、成体生物の系統発生に関連する胚の変化です。 進化の過程では、発達のすべての段階が再配置されます。 新しい変化は形成の最終段階で起こることがよくあります。 個体発生は段階を追加したり追加したりすることによってさらに複雑になります(同化)。 この場合にのみ、遠い先祖におけるこれらの部分の発達の歴史的段階を個体発生において繰り返すためのすべての前提条件が整います(反復)。 しかし、個体発生は、発達の他の段階で変化する可能性があり、その後のすべての段階が以前の経路から逸脱します (逸脱)。 最後に、器官や部分そのものの基本的な変化(アーチャラクシス)も考えられます。 その後、個体発生全体が変化し、子孫の個々の発達において、先祖の発達の歴史的段階の経過の順序を示すものがないことが判明します。
E. ヘッケルの生物遺伝の法則の生物学的本質
ヘッケルの生物遺伝法則とセベルツォフの系統発生理論は、形態学の発展と進化論そのものに重要な役割を果たしています。 動物の個々の発達に関する研究は、動物の歴史的発達を示す十分な証拠を提供しています。 生物遺伝の法則は、E. ヘッケルによって開発された三重並列処理の方法の重要な要素であり、これを利用して系統発生が再構築されます。 この方法は、形態学的、発生学的、古生物学的なデータの比較に基づいています。 系統発生を再構成する際、形態学者は今でもヘッケル原理を使用しており、これによれば、子孫の個体発生は祖先の系統発生の段階を一時的に繰り返し、再現するという基本的な生物遺伝の法則のみに頼って、進化のプロセスを説明することは不可能です。過ぎ去ったことを繰り返すこと自体が新しい何かを生み出すわけではありません。 地球上の生命は特定の生物の世代交代によって存在するため、その個体発生の変化によってその進化が起こります。 これらの変化は、特定の個体発生が祖先の形態によって定められた経路から逸脱し、新しい特徴を獲得するという事実に要約されます。
このような逸脱には、例えば、共新生、つまり胚や幼虫の中で生じ、それらを環境の特徴に適応させる適応が含まれます。 成体の生物では、新生は保存されません。 共生生殖の例としては、尾のない両生類の幼虫の口の中に角質が形成され、これにより幼虫は植物性食物を食べやすくなります。 カエルの変態の過程で、それらは消滅し、昆虫や虫を食べるために消化器系が再構築されます。 胎盤哺乳類とヒトの胎盤形成 - へその緒のある胎盤。
個体発生の初期段階にのみ現れるセノジェネシスは、成体生物の組織のタイプを変更しませんが、子孫の生存の可能性を高めます。 それらは、生殖能力の低下と胚または幼虫期間の延長を伴う可能性があり、これにより、胚後または幼虫後発育期の生物はより成熟して活動的になることが判明します。 セノジェネシスは発生し、有用であることが証明されたため、次の世代でも再現されるでしょう。
系統発生学的に重要な系統発生のもう 1 つのタイプの変換は、系統発生です。 それらは、祖先に特有の個体発生からの逸脱を表し、胚発生で明らかになりますが、成体の形態では適応的な重要性を持ちます。 したがって、哺乳動物では毛の形成は胚発生の非常に初期の段階で現れますが、毛自体が重要なのは成体の生物においてのみであり、個体発生におけるそのような変化は有用であり、自然選択によって固定され、次の世代で再生産されます。 これらの変化は、先天性奇形を引き起こすのと同じメカニズム、つまり細胞の増殖、移動、接着、死または分化の破壊に基づいていますが、新生と同様に、それらはその適応価値によって欠陥とは区別されます。 系統発生における自然選択による有用性と固定化。
胚形成と特定の構造の形態形成の段階に応じて、系統形成の重要性を持つ発生変化が発生し、3つのタイプが区別されます。
1. 同化作用または拡張は、器官の発達が実質的に完了した後に発生し、最終結果を変える追加の段階の追加として表現されます。 同化作用には、ヒラメが卵から孵化して稚魚になった後にのみ他の魚と見分けがつかない特定の体型を獲得する現象や、背骨の湾曲の出現、脳頭蓋骨の縫合糸の融合、最終的な変形などの現象が含まれます。哺乳類と人間の体内の血管の再分布。
2. 逸脱 - 器官の形態形成の過程で生じる逸脱。 例としては、哺乳類の個体発生における心臓の発達が挙げられます。この発達では、管段階、二室構造、三室構造が再現されていますが、爬虫類の特徴である不完全な中隔の形成段階は、心臓の発達段階に置き換えられています。隔壁の構造と位置が異なり、哺乳類にのみ特徴的です。 哺乳類の肺の発達では、祖先の初期段階の再現も見られ、その後の形態形成は新しい方法で進行します。
3. Archallaxis - 原始のレベルで検出され、それらの分割の違反、初期の分化、または根本的に新しい原基の出現として表現される変化。 アーカラキシの典型的な例は、哺乳類の毛の発達であり、その形成は発生の非常に初期段階で起こり、最初から他の脊椎動物の皮膚付属器の形成とは異なります。 弓頭蓋骨のタイプによると、脊索は原始的な頭蓋骨のない魚に現れ、軟骨脊椎は軟骨魚に現れ、二次腎臓のネフロンは爬虫類に発達します。
子孫の個体発生における同化作用による進化の過程で、基本的な生物遺伝の法則が完全に実現されることは明らかです。 祖先の発達段階すべてが再現されます。 逸脱の場合、初期の祖先段階が再現され、その後の段階は新しい方向への発展に置き換えられます。 Archallaxis は、これらの構造の発展における再現を完全に妨げ、その基本を変えてしまいます。
個体発生の進化において、同化は系統発生として最も頻繁に起こりますが、これは発生プロセス全体をほんのわずかしか変化させません。 胚形成における形態形成プロセスの違反としての逸脱は、多くの場合、自然選択によって拒否されるため、あまり一般的ではありません。 アーカラキシスは、胚形成の過程全体を変化させるという事実により、進化において最もまれに発生します。そして、そのような変化が重要な器官や、重要な胚組織の中心である器官の初歩に影響を与える場合、それらは生命と両立しないことが判明することがよくあります。
個体発生の進化においては、新生と系統発生に加えて、器官形成の時期の逸脱(異時性)やその発達の場所の逸脱(異所性)も見られます。 1つ目と2つ目の両方とも、発達中の構造の相互対応に変化をもたらし、自然選択の厳密な制御を受けます。 有用であることが判明した異時性と異性体のみが保存されます。 このような適応異時性の例としては、形成の種類に応じて進化する集団における最も重要な器官の形成時期の変化が挙げられます。 したがって、哺乳類、特に人間では、前脳の分化が他の部分の発達を著しく促進します。
ヘテロトピーは、器官間の新しい空間的および機能的接続の形成につながり、それらのさらなる共同進化を確実にします。 したがって、魚の咽頭の下に位置する心臓は、ガス交換のために鰓動脈への効率的な血液の流れを保証します。 陸生脊椎動物の胸骨後領域に移動すると、新しい呼吸器官である肺との単一複合体で発達して機能し、ここでまず第一に、ガス交換のために呼吸器系に血液を送達する機能を実行します。
ヘテロクロニーとヘテロトピーは、それらが胚形成と器官形態形成のどの段階で現れるかに応じて、異なるタイプの系統形成とみなすことができます。 ヒトの精巣が腹腔から鼠径管を通って陰嚢に至る異所性は、最終形成後の胚形成の終わりに観察され、典型的な同化です。
セノジェネシス、系統発生、異所性および異時性は、有用であることが証明されており、子孫において固定され、個体発生における新たな適応変化がそれらを置き換えて置き換えるまで、次の世代で再生産されます。 このおかげで、個体発生は私たちの祖先がたどった進化の道を一時的に繰り返すだけでなく、将来の系統発生の新しい方向性も切り開きます。