Mi a citoplazma felépítése és jelentősége. Citoplazma
Citoplazma(citoplazma) egy komplex kolloid rendszer, amely hialoplazmából, membránból és nem membrán organellumokból és zárványokból áll.
Hyaloplasma (a görög hialin szóból - átlátszó) egy összetett kolloid rendszer, amely különféle biopolimerekből (fehérjékből, nukleinsavakból, poliszacharidokból) áll, amely képes szolszerű (folyékony) állapotból gélbe és visszamenni.
¨A hialoplazma vízből, benne oldott szerves és szervetlen vegyületekből, valamint citomatrixból áll, amelyet 2-3 nm vastag fehérjeszálak trabekuláris hálója képvisel.
¨A hialoplazma funkciója az, hogy ez a közeg egyesíti az összes sejtszerkezetet, és biztosítja azok kémiai kölcsönhatását egymással.
A legtöbb intracelluláris transzportfolyamat a hialoplazmán keresztül megy végbe: aminosavak, zsírsavak, nukleotidok és cukrok átvitele. A hialoplazmában állandó ionáramlás folyik a plazmamembránba és onnan a mitokondriumokba, a sejtmagba és a vakuólumokba. A hialoplazma a citoplazma teljes térfogatának körülbelül 50% -át teszi ki.
Organellumok és zárványok. Az organellumok minden sejt számára állandó és kötelező mikrostruktúrák, amelyek biztosítják a létfontosságú sejtfunkciók ellátását.
Méretüktől függően az organellumokat a következőkre osztják:
1) mikroszkopikus - fénymikroszkóp alatt látható;
szubmikroszkópos - elektronmikroszkóppal megkülönböztethető.
A membrán jelenléte alapján az organellumok összetételében megkülönböztetik őket:
1) membrán;
nem membrán.
Céljuktól függően az összes organellum a következőkre oszlik:
Membránszervecskék
Mitokondriumok
A mitokondriumok általános célú mikroszkopikus membránszervecskék.
¨Méretek - vastagság 0,5 mikron, hossz 1-10 mikron.
¨ Alak - ovális, hosszúkás, szabálytalan.
¨Struktúra - a mitokondriumot két körülbelül 7 nm vastag membrán határolja:
1)Külső sima mitokondriális membrán(membrana mitochondrialis externa), amely elválasztja a mitokondriumot a hialoplazmától. Egyenlő körvonalú, és úgy van zárva, hogy egy zacskót képvisel.
Belső mitokondriális membrán(memrana mitochondrialis interna), amely a mitokondriumok belsejében kinövéseket, redőket (cristae) képez, és korlátozza a mitokondrium - a mátrix - belső tartalmát. A mitokondrium belsejét egy elektronsűrű anyag tölti ki, az ún
A mátrix finomszemcsés szerkezetű, 2-3 nm vastag vékony szálakat és körülbelül 15-20 nm méretű granulátumot tartalmaz. A szálak DNS-molekulák, a kis szemcsék pedig mitokondriális riboszómák.
¨ A mitokondriumok működése
1. Az energia szintézise és felhalmozódása ATP formájában a szerves szubsztrátok oxidációs folyamatai és az ATP foszforilációja eredményeként megy végbe. Ezek a reakciók a mátrixban lokalizált trikarbonsavciklus enzimek részvételével mennek végbe. A cristae membránokban további elektrontranszport és ehhez kapcsolódó oxidatív foszforiláció (ADP ATP-vé történő foszforilációja) rendszerei vannak.
2. Fehérjeszintézis. A mátrixukban lévő mitokondriumok autonóm fehérjeszintézis rendszerrel rendelkeznek. Ezek az egyetlen olyan organellák, amelyek saját, hisztonfehérjéktől mentes DNS-molekulákkal rendelkeznek. A mitokondriális mátrixban riboszómák képződnek is, amelyek számos olyan fehérjét szintetizálnak, amelyeket a sejtmag nem kódol, és saját enzimrendszerük felépítésére szolgál.
3. A vízanyagcsere szabályozása.
Lizoszómák
A lizoszómák (lizoszómák) általános célú szubmikroszkópos membránszervecskék.
¨Méretek - 0,2-0,4 mikron
¨ Alak - ovális, kicsi, gömb alakú.
¨Struktúra – a lizoszómák proteolitikus enzimeket tartalmaznak (több mint 60 ismert), amelyek képesek különböző biopolimerek lebontására. Az enzimek egy zárt membránzsákban helyezkednek el, ami megakadályozza, hogy bejussanak a hialoplazmába.
A lizoszómák négy típusa létezik:
Elsődleges lizoszómák;
Másodlagos (heterofagoszómák, fagolizoszómák);
Autofagoszómák
Maradék testek.
Elsődleges lizoszómák- ezek kisméretű, 0,2-0,5 µm méretű membránvezikulák, amelyek inaktív állapotban hidrolitikus enzimeket (marker - savas foszfatáz) tartalmazó, strukturálatlan anyaggal töltenek meg.
Másodlagos lizoszómák(heterofagoszómák) vagy intracelluláris emésztési vakuolák, amelyek primer lizoszómák és fagocita vakuolák fúziójával jönnek létre. Az elsődleges lizoszóma enzimei kapcsolatba lépnek a biopolimerekkel, és monomerekre bontják azokat. Ez utóbbiak a membránon keresztül a hialoplazmába kerülnek, ahol újra hasznosulnak, azaz részt vesznek a különböző anyagcsere-folyamatokban.
Autofagoszómák (autolizoszómák)– folyamatosan megtalálhatók a protozoonok, növények és állatok sejtjeiben. Morfológiájuk szerint a másodlagos lizoszómák közé sorolhatók, de azzal a különbséggel, hogy ezek a vakuolák fragmentumokat vagy akár teljes citoplazmatikus struktúrákat tartalmaznak, mint például mitokondriumok, plasztidok, riboszómák és glikogén granulátumok.
Maradék testek(telolysosome, corpusculum residuale) - biológiai membránnal körülvett, emésztetlen maradványok, kis mennyiségű hidrolitikus enzimet tartalmaznak, a tartalom tömörödik és átrendeződik. Gyakran a maradék testekben az emésztetlen lipidek másodlagos strukturálódása következik be, és ez utóbbiak réteges struktúrákat alkotnak. Pigmentanyagok lerakódása is előfordul - lipofuscint tartalmazó öregedő pigment.
¨Funkció - biogén makromolekulák emésztése, a sejt által szintetizált termékek módosítása hidrolázok segítségével.
Citoplazma - a sejt egy kötelező része, amely a plazmamembrán és a sejtmag közé záródik és képviseli hialoplazma - a citoplazma fő anyaga, organoidok- a citoplazma állandó komponensei és befogadás- a citoplazma átmeneti komponensei. A citoplazma kémiai összetétele változatos. Alapja víz (a citoplazma teljes tömegének 60-90%-a). A citoplazma fehérjében gazdag, tartalmazhat zsírokat és zsírszerű anyagokat, különféle szerves és szervetlen vegyületeket. A citoplazmában lúgos reakció van. A citoplazma egyik jellemző tulajdonsága az állandó mozgás (ciklózis). Elsősorban sejtszervecskék, például kloroplasztiszok mozgása révén észlelhető. Ha a citoplazma mozgása leáll, a sejt elpusztul, hiszen csak állandó mozgásban tudja ellátni funkcióit.
A citoplazma fő anyaga az hialoplazma (citoszol)- színtelen, nyálkás, sűrű és átlátszó kolloid oldat. Ebben zajlik le minden anyagcsere-folyamat, ez biztosítja a mag és az összes organellum összekapcsolódását. A folyékony rész vagy a nagy molekulák túlsúlyától függően a hialoplazmában a hialoplazma két formáját különböztetjük meg: sol - folyékonyabb hyaloplasma és gél- vastagabb hialoplazma. Kölcsönös átmenet lehetséges köztük: a gél könnyen szollá alakul és fordítva.
Sejtmembránok az eukarióta szervezetek eltérő felépítésűek, de a plazmamembrán mindig a citoplazmával szomszédos, és a felületén külső réteg képződik. Az állatoknál úgy hívják glikokalix(glikoproteinek, glikolipidek, lipoproteinek alkotják), növényekben - sejtfal vastag rostrétegből.
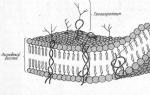
Membrán szerkezet. Minden biológiai membránnak közös szerkezeti jellemzői és tulajdonságai vannak. Jelenleg általánosan elfogadott folyékony mozaik modell membránszerkezet (szendvics modell). A membrán alapvetően egy lipid kettős rétegen alapul foszfolipidek. Egy kettős rétegben a membránban lévő molekulák farka egymással szemben, a poláris fejek pedig kifelé, a víz felé néznek. A lipideken kívül a membrán fehérjéket is tartalmaz (átlagosan 60%). Ezek határozzák meg a membrán legtöbb specifikus funkcióját. A fehérjemolekulák nem alkotnak folyamatos réteget; perifériás fehérjék- a lipid kettősréteg külső vagy belső felületén található fehérjék, félig integrált fehérjék- a lipid kettősrétegben változó mélységig elmerülő fehérjék, integrál, vagy transzmembrán fehérjék- fehérjék, amelyek áthatolnak a membránon, érintkezve a sejt külső és belső környezetével egyaránt.
A membránfehérjék többféle funkciót is elláthatnak: bizonyos molekulák transzportja, a membránokon végbemenő reakciók katalizálása, a membránok szerkezetének fenntartása, a környezetből érkező jelek fogadása és átalakítása.
A membrán 2-10% szénhidrátot tartalmazhat. A membránok szénhidrát komponensét általában fehérjemolekulákhoz (glikoproteinekhez) vagy lipidekhez (glikolipidekhez) kapcsolódó oligoszacharid vagy poliszacharid láncok képviselik. A szénhidrátok főként a membrán külső felületén helyezkednek el. A szénhidrátok biztosítják a membrán receptor funkcióit. Az állati sejtekben a glikoproteinek egy membrán feletti komplexet alkotnak - glikokalix, vastagsága több tíz nanométer. Extracelluláris emésztés megy végbe benne, sok sejtreceptor található, és a sejtadhézió láthatóan a segítségével megy végbe.
A fehérjék és lipidek molekulái mozgékonyak, elsősorban a membrán síkjában képesek mozogni. A plazmamembrán vastagsága átlagosan 7,5 nm.
Membrán funkciók.
1. Elválasztják a sejttartalmat a külső környezettől.
2. Szabályozza a sejt és a környezet közötti anyagcserét.
3. A sejteket kompartmentekre osztják, amelyeket különféle reakciók lebonyolítására terveztek.
4. Számos kémiai reakció megy végbe magán a membránokon elhelyezett enzimatikus szállítószalagokon.
5. Biztosítson kommunikációt a sejtek között a többsejtű élőlények szöveteiben.
6. A külső ingerek felismerésére szolgáló receptorterületek a membránokon helyezkednek el.
A membrán egyik fő funkciója a szállítás, amely biztosítja az anyagcserét a sejt és a külső környezet között. A membránoknak megvan a tulajdonuk szelektív permeabilitás, azaz egyes anyagok vagy molekulák számára jól áteresztők, mások számára pedig rosszul (vagy teljesen áthatolhatatlanok). Különféle mechanizmusok léteznek az anyagoknak a membránon keresztül történő szállítására. Attól függően, hogy az anyagok szállításához energiafelhasználás szükséges, megkülönböztetik őket : passzív szállítás- anyagok szállítása energiafogyasztás nélkül; aktív közlekedés - energiát igénylő közlekedés.
A passzív transzport a koncentrációk és a töltések különbségén alapul. A passzív szállítás során az anyagok mindig a magasabb koncentrációjú területről egy alacsonyabb koncentrációjú területre, azaz koncentrációgradiens mentén mozognak.
Megkülönböztetni A passzív transzport három fő mechanizmusa:egyszerű diffúzió- anyagok szállítása közvetlenül a lipid kettős rétegen keresztül. Gázok, nem poláris vagy kis töltés nélküli poláris molekulák könnyen átjutnak rajta. Minél kisebb a molekula és minél jobban oldódik zsírban, annál gyorsabban hatol be a membránon. Érdekes módon a poláris vízmolekulák nagyon gyorsan behatolnak a lipid kettős rétegbe. Ez azzal magyarázható, hogy molekulái kicsiek és elektromosan semlegesek. A víz membránokon keresztül történő diffúzióját ún ozmózissal.
Diffúzió membráncsatornákon keresztül. A töltött molekulák és ionok (Na +, K +, Ca 2+, C1~) egyszerű diffúzióval nem képesek átjutni a lipid kettősrétegen, azonban a pórusokat alkotó speciális csatornaképző fehérjék jelenléte miatt áthatolnak a membránon. . A víz nagy része az akvaporin fehérjék által kialakított csatornákon keresztül halad át a membránon.
Könnyített diffúzió- anyagok szállítása speciális transzportfehérjék segítségével, amelyek mindegyike felelős bizonyos molekulák vagy rokon molekulacsoportok szállításáért. Kölcsönhatásba lépnek a szállított anyag egy molekulájával, és valahogy áthelyezik a membránon. Így kerül a sejtbe a cukrok, aminosavak, nukleotidok és sok más poláris molekula.
Szükség aktív közlekedés akkor fordul elő, amikor biztosítani kell a molekulák transzportját a membránon az elektrokémiai gradiens ellen. Ezt a transzportot hordozófehérjék végzik, amelyek tevékenységéhez energiára van szükség. Az energiaforrás az ATP molekulák. Az egyik legtöbbet tanulmányozott aktív transzportrendszer a nátrium-kálium pumpa. A K + koncentrációja a sejten belül sokkal magasabb, mint azon kívül, és a Na + - fordítva. Ezért a K + a membrán vízpórusain keresztül passzívan kidiffundál a sejtből, a Na + pedig a sejtbe. Ugyanakkor a sejt normális működéséhez fontos a K + és Na + ionok bizonyos arányának fenntartása a citoplazmában és a külső környezetben. Ez azért lehetséges, mert a membrán a nátrium-kálium pumpának köszönhetően aktívan pumpálja a Na +-t a sejtből és a K +-t a sejtbe. A sejtek működéséhez szükséges energia csaknem egyharmadát a nátrium-kálium pumpa működésére fordítják. Egy működési ciklus során a szivattyú 3 Na + iont pumpál ki a cellából és 2 K + iont pumpál be. A K+ passzívan gyorsabban diffundál ki a sejtből, mint a Na+ a sejtbe.
A sejtnek olyan mechanizmusai vannak, amelyek révén nagy részecskéket és makromolekulákat tud szállítani a membránon keresztül. A makromolekulák sejt általi felszívódásának folyamatát ún endocitózis. Az endocitózis során a plazmamembrán invaginációt képez, szélei összeolvadnak, és a citoplazmából egyetlen membrán határol el struktúrákat, amelyek a külső citoplazmatikus membrán részét képezik. Az endocitózisnak két típusa van: fagocitózis- nagy részecskék befogása és abszorpciója (például limfociták, protozoák fagocitózisa stb.) és pinocitózis - folyadékcseppek felfogásának és felszívásának folyamata a benne oldott anyagokkal.
Exocitózis- a különböző anyagok sejtből történő eltávolításának folyamata. Az exocitózis során a hólyagmembrán egyesül a külső citoplazmatikus membránnal, a vezikula tartalma a sejten kívülre kerül, membránja pedig a külső citoplazmatikus membránba kerül.
Sejtorganellumok
Organellumok (organellumok)- állandó sejtszerkezetek, amelyek biztosítják a sejt meghatározott funkciók ellátását. Minden organellumnak sajátos szerkezete van, és meghatározott funkciókat lát el.
Vannak: membránszervecskék - membránszerkezettel rendelkeznek, és lehetnek egymembránosak (endoplazmatikus retikulum, Golgi-készülék, lizoszómák, növényi sejtek vakuólumai) és kettős membránok (mitokondriumok, plasztidok, mag).
A membránon kívül lehetnek nem membrán organellák is - olyanok, amelyeknek nincs membránszerkezete (kromoszómák, riboszómák, sejtközpont és centriolok, csillók és flagellák alaptesttel, mikrotubulusok, mikrofilamentumok).
Egymembrán organellumok:
1. Endoplazmatikus retikulum (ER). Ez egy olyan membránrendszer, amely tartályokat és csatornákat képez, amelyek egymáshoz kapcsolódnak és egyetlen belső teret határolnak le - EPR üregek. A membránok egyrészt a külső citoplazmatikus membránhoz, másrészt a nukleáris membrán külső héjához kapcsolódnak. Kétféle EPR létezik: durva (szemcsés), felületén riboszómákat tartalmaz, és lapított zsákok gyűjteményét képviseli, és sima (agranuláris), amelyek membránjai nem hordoznak riboszómákat.
Funkciói: a sejt citoplazmáját izolált kompartmentekre osztja, ezáltal biztosítva a számos, párhuzamosan lezajló reakció egymástól való térbeli elhatárolását, elvégzi a szénhidrátok és lipidek szintézisét és lebontását (sima ER) és biztosítja a fehérjeszintézist (durva ER), csatornákban és üregekben halmozódik fel, majd bioszintetikus termékeket szállít a sejtszervecskékbe.
2. Golgi-készülék.Általában a sejtmag közelében (állati sejtekben, gyakran a sejtközpont közelében) található organellum. Kiszélesedett szélű, lapított ciszternák halmaza, amelyhez kis egymembrános vezikulák (Golgi-vezikulák) rendszere társul. Minden köteg általában 4-6 tartályból áll. A Golgi-veremek száma egy cellában egytől több százig terjed.
A Golgi komplexum legfontosabb funkciója a különféle váladékok (enzimek, hormonok) eltávolítása a sejtből, ezért jól fejlett a kiválasztó sejtekben. Itt az összetett szénhidrátok szintézise egyszerű cukrokból, a fehérjék érése és a lizoszómák képződése következik be.
3. Lizoszómák. A legkisebb egymembrán sejtszervecskék, amelyek 0,2-0,8 mikron átmérőjű vezikulák, amelyek legfeljebb 60 hidrolitikus enzimet tartalmaznak, amelyek enyhén savas környezetben működnek.
A lizoszómák képződése a Golgi apparátusban megy végbe, ahová az ER-ből érkeznek a benne szintetizált enzimek. Az anyagok enzimek segítségével történő lebontását lízisnek nevezik, innen ered az organellum neve.
Vannak: primer lizoszómák - a Golgi-apparátusról leválasztott és inaktív formában lévő enzimeket tartalmazó lizoszómák, valamint másodlagos lizoszómák - primer lizoszómák pinocitotikus vagy fagocitotikus vakuolákkal való fúzió eredményeként képződő lizoszómák; A sejtbe jutó anyagok emésztése, lízise megy végbe bennük (ezért szokták emésztési vakuólumoknak is nevezni).
Az emésztés termékeit a sejt citoplazmája szívja fel, de az anyag egy része emésztetlenül marad. Az ezt az emésztetlen anyagot tartalmazó másodlagos lizoszómát maradéktestnek nevezzük. Exocitózissal az emésztetlen részecskéket eltávolítják a sejtből.
Néha a sejt önmegsemmisítése lizoszómák részvételével történik. Ezt a folyamatot autolízisnek nevezik. Ez általában bizonyos differenciálódási folyamatok során fordul elő (például a porcos szövet csontszövettel való helyettesítése, a béka ebihalnál a farok eltűnése).
4. Cilia és flagella. Kilenc kettős mikrotubulus alkotja, amelyek egy membránnal borított henger falát alkotják; közepén két egyetlen mikrotubulus található. Ez a 9+2 típusú szerkezet a csillókra és flagellákra jellemző szinte minden eukarióta szervezetben, a protozoontól az emberig.
A csillókat és a flagellákat a citoplazmában erősítik az ezeknek az organellumoknak a tövében elhelyezkedő bazális testek. Mindegyik alaptest kilenc mikrotubulusból áll, a közepén nincsenek mikrotubulusok.
5. Az egymembrán organellumok közé tartoznak még vakuolák, membrán veszi körül - tonoplaszt. A növényi sejtekben a sejttérfogat 90%-át is elfoglalhatják, és a magas ozmotikus potenciál és turgor (intracelluláris nyomás) révén biztosítják a víz beáramlását a sejtbe. Az állati sejtekben a vakuolák kicsik, endocitózis (fagocitotikus és pinocitotikus) következtében alakulnak ki, a primer lizoszómákkal való egyesülés után emésztési vakuólumoknak nevezik őket.
Kettős membrán organellumok:
1. Mitokondriumok. Eukarióta sejt kettős membrán organellumai, amelyek energiával látják el a szervezetet. A sejtben található mitokondriumok száma tág határok között változik, 1-100 ezer között, és az anyagcsere aktivitásától függ. A mitokondriumok száma osztódással növekedhet, mivel ezeknek az organellumoknak saját DNS-ük van.
A mitokondriumok külső membránja sima, a belső membrán számos invaginációt vagy tubuláris kinövést képez - cristas. A cristae száma a sejt funkcióitól függően több tíztől több százig, sőt ezerig is változhat. Megnövelik a belső membrán felületét, amelyen az ATP molekulák szintézisében részt vevő enzimrendszerek találhatók.
A mitokondriumok belső tere megtelt mátrix. A mátrix egy körkörös mitokondriális DNS-molekulát, specifikus mRNS-t, tRNS-t és riboszómákat (prokarióta típus) tartalmaz, amelyek a belső membránt alkotó fehérjék egy részének autonóm bioszintézisét végzik. Ezek a tények alátámasztják a mitokondriumok oxidáló baktériumokból való eredetét (a szimbiogenezis hipotézis szerint). De a legtöbb mitokondriális gén a sejtmagba költözött, és számos mitokondriális fehérje szintézise a citoplazmában megy végbe. Ezenkívül olyan enzimeket tartalmaz, amelyek ATP-molekulákat képeznek. A mitokondriumok hasadással képesek szaporodni.
A mitokondriumok funkciói a szénhidrátok, aminosavak, glicerin és zsírsavak oxigénbontása ATP képződésével, a mitokondriális fehérjék szintézise.
2. Plasztidok. A plasztidoknak három fő típusa van: leukoplasztok- színtelen plasztidok a színtelen növényrészek sejtjeiben, kromoplasztok- színes plasztidok, általában sárga, piros és narancssárga, kloroplasztiszok- zöld plasztiszok. A plasztidok proplasztidokból - legfeljebb 1 mikron méretű kettős membrános vezikulákból - keletkeznek.
Mivel a plasztidok közös eredetûek, lehetséges köztük a kölcsönös átalakulás. Leggyakrabban a leukoplasztok kloroplasztokká alakulnak (a burgonyagumók fényben zöldítése a sötétben történik). Amikor a levelek sárgulnak és a gyümölcsök pirosra fordulnak, a kloroplasztok kromoplasztokká alakulnak. Csak a kromoplasztok leukoplasztokká vagy kloroplasztokká történő átalakulását tartják lehetetlennek.
Kloroplasztok. A fő funkció a fotoszintézis, azaz. A kloroplasztiszokban a napenergia ATP-molekulák energiájává történő átalakítása következtében szerves anyagok szintetizálódnak a szervetlenekből. A magasabb rendű növények kloroplasztiszai bikonvex lencse alakúak. A külső membrán sima, a belső pedig hajtogatott szerkezetű. A belső membrán kiemelkedéseinek kialakulása következtében lamellák és tilakoidok rendszere keletkezik. A kloroplasztiszok belső környezete - stroma körkörös DNS-t és prokarióta típusú riboszómákat tartalmaz. A plasztidok képesek autonóm osztódásra, mint a mitokondriumok. A szimbiogenezis hipotézise szerint bizonyítékok is alátámasztják a plasztidok cianobaktériumokból való eredetét.
Rizs. A növényi sejt szerkezetének modern (általánosított) diagramja, különböző növényi sejtek elektronmikroszkópos vizsgálata alapján összeállított: 1 - Golgi készülék; 2 - szabadon elhelyezkedő riboszómák; 3 - kloroplasztiszok; 4 - intercelluláris terek; 5 - poliriboszómák (több riboszóma összekapcsolódik); 6 - mitokondriumok; 7 - lizoszómák; 8 - szemcsés endoplazmatikus retikulum; 9 - sima endoplazmatikus retikulum; 10 - mikrotubulusok; 11 - plasztiszok; 12 - a membránon áthaladó plazmodezma; 13 - sejtmembrán; 14 - nucleolus; 15, 18 - nukleáris burok; 16 - pórusok a nukleáris burokban; 17 - plazmalemma; 19 - hialoplazma; 20 - tonoplaszt; 21 - vakuolák; 22 - mag.

Rizs. Membrán szerkezet

Rizs. A mitokondriumok szerkezete. Felül és középen - egy mitokondriumon keresztüli hosszmetszet nézete (fent - egy mitokondrium a gyökércsúcs embrionális sejtjéből; középen - egy felnőtt elodea levél sejtjéből). Az alábbiakban látható egy háromdimenziós diagram, amelyen a mitokondriumok egy része le van vágva, ami lehetővé teszi a belső szerkezetének megtekintését. 1 - külső membrán; 2 - belső membrán; 3 - cristae; 4 - mátrix.

Rizs. Kloroplaszt szerkezet. A bal oldalon - a kloroplaszton áthaladó hosszanti metszet: 1 - halomba hajtogatott lamellák által alkotott grana; 2 - héj; 3 - stroma (mátrix); 4 - lamellák; 5 - csepp zsír képződik a kloroplasztiszban. A jobb oldalon egy háromdimenziós diagram a lamellák és a gránák elhelyezkedéséről és kapcsolatáról a kloroplaszton belül: 1 - grana; 2 - lamellák.
2. A citoplazma szerkezete, kémiai összetétele, jelentősége. A membránok felépítése és funkciói.
Citoplazma (protoplazma) a sejt élő tartalmát már a 12. században ismerték. A protoplazma kifejezést először Purkinje cseh tudós javasolta (1839).
A citoplazmának három rétege van: plazmalemma, hialoplazma és tonoplaszt.
Plasmalemma - elemi membrán, a citoplazma külső rétege, a membrán mellett. Vastagsága körülbelül 80A (A - angström, 10-10 m). Foszfolipidekből, fehérjékből, lipoproteinekből, szénhidrátokból, szervetlen ionokból, vízből áll. Lehet lamellás (réteges) és micellás (csepp) szerkezetű. Leggyakrabban 3 rétegből áll: foszfolipidek bimolekuláris rétege (35A), részarányuk 40%, felületét mindkét oldalon nem folytonos szerkezeti fehérjék (20 és 25A) borítják. Egyes helyeken a lamelláris és micelláris struktúrák találkozásánál vagy két micella között a szerkezeti fehérjék külső és belső rétege összezáródhat, hidrofil fehérjepórusokat, 7-10A-t képezve, amelyeken az anyagok oldott állapotban haladnak át.
Az enzimatikus aktivitással nem rendelkező fehérjemolekulák beépülnek a membránmátrixba - specifikus szelektív ionvezetési csatornák (kálium, nátrium stb.). Végül a membrán fehérjéket tartalmazhat - olyan enzimeket, amelyek biztosítják a nagy molekulájú anyagok bejutását a sejtbe. Mindezek a képződmények - biokémiai pórusok - biztosítják a membránok fő tulajdonságát - a félig áteresztő képességet.
A plazmalemmának számos redő, mélyedés és kiemelkedés van, ami sokszorosára növeli a felületét.
A plazmalemma membránként fontos és összetett funkciókat lát el: 1. Szabályozza a sejt általi anyagok felvételét és kibocsátását; 2. Átalakítja, tárolja és fogyasztja az energiát; 3. kémiai átalakítót képvisel, felgyorsítja az anyagok átalakulását; 4. Fogadja és átalakítja a külvilág fény-, mechanikai és kémiai jeleit.
Így a plazmalemma szabályozza a sejt permeabilitását, az anyagok felszívódási, átalakulási, szekréciós és kiválasztási folyamatait.
Tonoplast - belső membrán, amely elválasztja a sejtnedvet a citoplazmától
Hialoplazma. A sejtszerveződés alapját képviseli, élőlényként kifejezi a lényegét. Fizikai-kémiai szempontból összetett heterogén kolloid rendszer, ahol nagy molekulatömegű vegyületek vizes környezetben diszpergálódnak. A citoplazma átlagosan 70-80% vizet, 12% fehérjét, 1,5-2% nukleinsavat, körülbelül 5% zsírt, 4-6% szénhidrátot és 0,5-2% szervetlen anyagot tartalmaz. Két halmazállapotú lehet: szol és gél. Sol- folyékony halmazállapotú, viszkozitása van, gél- szilárd halmazállapotú, rugalmas, nyújtható. Képes reverzibilis szol-gél átmenetekre hőmérséklet, hidrogénion-koncentráció, elektrolit hozzáadása és mechanikai hatás hatására.
Citoplazmaállandóban van mozgás, ami normál körülmények között nagyon lassú és szinte észrevehetetlen. A hőmérséklet emelkedése, fény- vagy kémiai inger felgyorsítja a citoplazma mozgását, és fénymikroszkópban láthatóvá teszi. A kloroplasztok, amelyeket a viszkózus citoplazma árama visz el, segítik ezt a mozgást. A citoplazma mozgása kétféle: körkörös (forgó) és csíkos (keringési). Ha a sejtüreget egy nagy vakuólum foglalja el, akkor a citoplazma csak a falak mentén mozog. Ez egy körkörös mozgás. Megfigyelhető a Vallisneria és az Elodea levélsejtjeiben. Ha egy sejtben több vakuólum van, akkor a sejten keresztező citoplazma szálai a sejtmag központjában kapcsolódnak egymáshoz. Ezekben a zsinórokban a citoplazma csíkos mozgása történik. A csalán csípős szőrének sejtjeiben és a tök fiatal hajtásainak szőrsejtjeiben a citoplazma csíkos mozgása figyelhető meg.
A hialoplazma tulajdonságai a fehérjetermészetű szupramolekuláris struktúrákhoz is kapcsolódnak. Ezek mikrotubulusok és mikrofilamentumok.
Mikrotubulusok- üreges kis képződmények elektronsűrű fehérjefallal. Részt vesznek az anyagok citoplazmán történő vezetésében, a kromoszómák mozgásában és a mitotikus orsószálak kialakításában.
Mikrofilamentumok spirálisan elrendezett fehérje alegységekből állnak, amelyek rostokat vagy háromdimenziós hálózatot alkotnak, kontraktilis fehérjéket tartalmaznak, és elősegítik a hialoplazma és a hozzájuk kapcsolódó organellumok mozgását.
Hyaloplasma mint makromolekulák és szupramolekuláris struktúrák komplex heterogén kolloid rendszere, vízben való oldhatatlansága, viszkozitása, rugalmassága, fordított változási képesség, a természetes membránok pórusain való átjárhatatlanság, nagy határfelületek, erős fénytörés és nagyon alacsony diffúzió jellemzi. arány.
Hyaloplasma organellumok . Amint korábban megjegyeztük, a hialoplazma nagyszámú szupramolekuláris képződményt tartalmaz, amelyek számos organellát képviselnek.
A biomembránok funkciói
1) gát – szabályozott, szelektív, passzív és aktív anyagcserét biztosít a környezettel. Például a peroxiszóma membrán megvédi a citoplazmát a sejtre veszélyes peroxidoktól. A szelektív permeabilitás azt jelenti, hogy a membrán különböző atomok vagy molekulák permeabilitása azok méretétől, elektromos töltésétől és kémiai tulajdonságaitól függ. A szelektív permeabilitás biztosítja, hogy a sejt és a sejtrekesz elkülönüljön a környezettől és el legyen látva a szükséges anyagokkal.
2) transzport – az anyagok a membránon keresztül jutnak be a sejtbe és onnan ki. A membránokon keresztül történő szállítás biztosítja: a tápanyagok szállítását, az anyagcsere végtermékeinek eltávolítását, a különböző anyagok kiválasztását, az iongradiensek létrehozását, a megfelelő pH és ionkoncentráció fenntartását a sejtben, amelyek a sejtenzimek működéséhez szükségesek. bármilyen okból nem képesek átjutni a foszfolipid kettősrétegen (például hidrofil tulajdonságai miatt, mivel a belsejében lévő membrán hidrofób és nem engedi át a hidrofil anyagokat, vagy nagy mérete miatt), de szükséges a sejt számára , speciális hordozófehérjéken (transzportereken) és csatornafehérjéken keresztül, illetve endocitózissal átjuthatnak a membránon. Ennek a mechanizmusnak egy változata a megkönnyített diffúzió, amelyben egy adott molekula segíti az anyagot átjutni a membránon. Ennek a molekulának lehet egy csatornája, amelyen csak egyfajta anyag jut át. Az aktív transzport energiát igényel, mivel a koncentráció gradienssel szemben történik. A membránon speciális pumpás fehérjék találhatók, köztük az ATPáz, amelyek aktívan pumpálják a káliumionokat (K+) a sejtbe, és nátriumionokat (Na+) pumpálnak ki belőle.
3) mátrix - biztosítja a membránfehérjék bizonyos relatív helyzetét és orientációját, optimális kölcsönhatásukat;
4) mechanikus - biztosítja a sejt autonómiáját, intracelluláris struktúráit, valamint a kapcsolatot más sejtekkel (a szövetekben). A sejtfalak nagy szerepet játszanak a mechanikai működés biztosításában, állatoknál pedig az intercelluláris anyag.
5) energia - a kloroplasztiszokban a fotoszintézis és a mitokondriumokban a sejtlégzés során a membránjukban energiatranszfer rendszerek működnek, amelyekben fehérjék is részt vesznek;
6) receptor - a membránban található egyes fehérjék receptorok (molekulák, amelyek segítségével a sejt bizonyos jeleket érzékel például a vérben keringő hormonok csak azokra a célsejtekre hatnak, amelyeknek ezeknek a hormonoknak megfelelő receptorai vannak). A neurotranszmitterek (az idegimpulzusok vezetését biztosító vegyszerek) speciális receptorfehérjékhez is kötődnek a célsejtekben.
7) enzimatikus - a membránfehérjék gyakran enzimek. Például a bélhámsejtek plazmamembránjai emésztőenzimeket tartalmaznak.
8) a biopotenciálok generálásának és vezetésének megvalósítása.
A membrán segítségével állandó ionkoncentrációt tartanak fenn a sejtben: a sejten belüli K+-ion koncentrációja sokkal magasabb, mint a külső, a Na+-koncentráció pedig jóval alacsonyabb, ami nagyon fontos, hiszen ez biztosítja a potenciálkülönbség fenntartása a membránon és az idegimpulzus generálása.
9) sejtjelölés - a membránon antigének vannak, amelyek markerként működnek - „címkék”, amelyek lehetővé teszik a sejt azonosítását. Ezek glikoproteinek (vagyis olyan fehérjék, amelyekhez elágazó oligoszacharid oldalláncok kapcsolódnak), amelyek az „antennák” szerepét töltik be. Az oldalláncok számtalan konfigurációja miatt lehetséges, hogy minden sejttípushoz specifikus markert készítsünk. A markerek segítségével a sejtek felismerhetnek más sejteket, és velük együtt tudnak működni, például szervek és szövetek kialakításában. Ez azt is lehetővé teszi, hogy az immunrendszer felismerje az idegen antigéneket.
A sejt membránnal határolt gélszerű tartalmát az élő sejt citoplazmájának nevezzük. A koncepciót 1882-ben Eduard Strassburger német botanikus vezette be.
Szerkezet
A citoplazma bármely sejt belső környezete, és a baktériumok, növények, gombák és állatok sejtjeire jellemző.
A citoplazma a következő összetevőkből áll:
- hialoplazma (citoszolok) - folyékony anyag;
- sejtzárványok - a sejt opcionális komponensei;
- organellumok - a sejt állandó összetevői;
- citoszkeleton - sejtváz.
A citoszol kémiai összetétele a következő anyagokat tartalmazza:
- víz - 85%;
- fehérjék - 10%
- szerves vegyületek - 5%.
A szerves vegyületek közé tartoznak:
- ásványi sók;
- szénhidrátok;
- lipidek;
- nitrogéntartalmú vegyületek;
- kis mennyiségű DNS és RNS;
- glikogén (az állati sejtekre jellemző).

Rizs. 1. A citoplazma összetétele.
A citoplazma tápanyagokat (zsírcseppeket, poliszacharidszemcséket), valamint a sejt oldhatatlan salakanyagait tartalmazza.
A citoplazma színtelen, folyamatosan mozog és áramlik. Tartalmazza a sejt összes organellumát, és közvetíti azok összekapcsolódását. Részleges eltávolításakor a citoplazma helyreáll. Amikor a citoplazmát teljesen eltávolítják, a sejt elpusztul.
A citoplazma szerkezete heterogén. Feltételesen kiosztani két citoplazmaréteg:
TOP 4 cikkakik ezzel együtt olvasnak
- ektoplazma (plazmagel) - külső sűrű réteg, amely nem tartalmaz organellákat;
- endoplazma (plazmaszol) - belső, folyékonyabb réteg, amely organellumokat tartalmaz.
Az ektoplazmára és endoplazmára való felosztás egyértelműen kifejeződik a protozoonokban. Az ektoplazma segíti a sejt mozgását.
Kívül a citoplazmát citoplazmatikus membrán vagy plazmalemma veszi körül. Megvédi a sejtet a károsodástól, szelektív anyagszállítást végez, és biztosítja a sejt ingerlékenységét. A membrán lipidekből és fehérjékből áll.
Élettevékenység
A citoplazma létfontosságú anyag, amely részt vesz a sejt fő folyamataiban:
- anyagcsere;
- növekedés;
- osztály.
A citoplazma mozgását ciklózisnak vagy citoplazmatikus áramlásnak nevezik. Eukarióta sejtekben fordul elő, beleértve az embert is. A ciklózis során a citoplazma anyagokat szállít a sejt összes organellumába, és a sejtanyagcserét végzi. A citoplazma az ATP fogyasztásával mozog a citoszkeletonon.
A citoplazma térfogatának növekedésével a sejt növekszik. Az eukarióta sejt testének a magosztódást követő osztódási folyamatát (kariokinézis) citokinézisnek nevezik. A testosztódás eredményeként a citoplazma az organellumokkal együtt két leánysejt között oszlik el.

Rizs. 2. Citokinézis.
Funkciók
A sejtben a citoplazma fő funkcióit a táblázat írja le.
A citoplazmának a membrántól való elválasztását a kilépő víz ozmózisával plazmolízisnek nevezzük. A fordított folyamat - deplazmolízis - akkor következik be, amikor elegendő mennyiségű víz kerül a sejtbe. A folyamatok minden sejtre jellemzőek, kivéve az állati sejteket.
Citoplazma- a sejt kötelező része, a plazmamembrán és a sejtmag közé zárva; hialoplazmára (a citoplazma fő anyaga), organellumokra (a citoplazma állandó összetevői) és zárványokra (a citoplazma ideiglenes összetevőire) oszlik. A citoplazma kémiai összetétele: az alap a víz (a citoplazma teljes tömegének 60-90%-a), különféle szerves és szervetlen vegyületek. A citoplazmában lúgos reakció van. Az eukarióta sejt citoplazmájának jellemző sajátossága az állandó mozgás ( ciklózis). Elsősorban sejtszervecskék, például kloroplasztiszok mozgása révén észlelhető. Ha a citoplazma mozgása leáll, a sejt elpusztul, hiszen csak állandó mozgásban tudja ellátni funkcióit.
Hyaloplasma ( citoszol) színtelen, nyálkás, vastag és átlátszó kolloid oldat. Ebben zajlik le minden anyagcsere-folyamat, ez biztosítja a mag és az összes organellum összekapcsolódását. A folyékony rész vagy a nagy molekulák túlsúlyától függően a hialoplazmában a hialoplazma két formáját különböztetjük meg: sol- folyékonyabb hyaloplasma és gél- vastagabb hialoplazma. Kölcsönös átmenet lehetséges köztük: a gél szollá alakul és fordítva.
A citoplazma funkciói:
- az összes sejtkomponens egyetlen rendszerbe való egyesítése,
- környezet számos biokémiai és élettani folyamat lefolyásához,
- környezet az organellumok létezéséhez és működéséhez.
Sejtmembránok
Sejtmembránok korlátozza az eukarióta sejteket. Minden sejtmembránban legalább két réteget lehet megkülönböztetni. A belső réteg a citoplazmával szomszédos, és a jelölése plazmamembrán(szinonimák - plazmalemma, sejtmembrán, citoplazmatikus membrán), amely felett kialakul a külső réteg. Állati sejtben vékony és ún glikokalix(glikoproteinek, glikolipidek, lipoproteinek alkotják), növényi sejtben - vastag, ún. sejtfal(cellulóz alkotja).
Minden biológiai membránnak közös szerkezeti jellemzői és tulajdonságai vannak. Jelenleg általánosan elfogadott membránszerkezet folyadékmozaik modellje. A membrán alapja egy lipid kettős réteg, amelyet főleg foszfolipidek alkotnak. A foszfolipidek olyan trigliceridek, amelyekben az egyik zsírsavmaradékot foszforsavmaradék helyettesíti; A molekula foszforsavmaradékot tartalmazó szakaszát hidrofil fejnek, a zsírsavmaradékot tartalmazó szakaszokat hidrofób faroknak nevezzük. A membránban a foszfolipidek szigorúan rendezetten helyezkednek el: a molekulák hidrofób farkai egymással szemben, a hidrofil fejek pedig kifelé, a víz felé néznek.
A lipideken kívül a membrán fehérjéket is tartalmaz (átlagosan ≈ 60%). Ezek határozzák meg a membrán specifikus funkcióinak nagy részét (bizonyos molekulák transzportja, reakciók katalízise, a környezetből érkező jelek fogadása és átalakítása stb.). Vannak: 1) perifériás fehérjék(a lipid kettős réteg külső vagy belső felületén található), 2) félig integrált fehérjék(változó mélységig a lipid kettős rétegbe merülve), 3) integrált vagy transzmembrán fehérjék(átszúrja a membránt, érintkezve a sejt külső és belső környezetével is). Az integrál fehérjéket bizonyos esetekben csatornaképző vagy csatornafehérjéknek nevezik, mivel hidrofil csatornáknak tekinthetők, amelyeken keresztül a poláris molekulák bejutnak a sejtbe (a membrán lipidkomponense nem engedi át őket).

A - hidrofil foszfolipid fej; B - hidrofób foszfolipid farok; 1 - az E és F fehérjék hidrofób régiói; 2 – az F fehérje hidrofil régiói; 3 - elágazó oligoszacharid lánc, amely egy lipidhez kapcsolódik egy glikolipid molekulában (a glikolipidek ritkábban fordulnak elő, mint a glikoproteinek); 4 - elágazó oligoszacharid lánc, amely egy fehérjéhez kapcsolódik egy glikoprotein molekulában; 5 - hidrofil csatorna (pórusként működik, amelyen keresztül az ionok és néhány poláris molekula átjuthat).
A membrán szénhidrátot tartalmazhat (legfeljebb 10%). A membránok szénhidrát komponensét fehérjemolekulákhoz (glikoproteinekhez) vagy lipidekhez (glikolipidekhez) kapcsolódó oligoszacharid vagy poliszacharid láncok képviselik. A szénhidrátok főként a membrán külső felületén helyezkednek el. A szénhidrátok biztosítják a membrán receptor funkcióit. Az állati sejtekben a glikoproteinek egy membrán feletti komplexet, a glikokalixot alkotnak, amely több tíz nanométer vastag. Sok sejtreceptort tartalmaz, segítségével sejtadhézió jön létre.
A fehérjék, szénhidrátok és lipidek molekulái mozgékonyak, képesek a membrán síkjában mozogni. A plazmamembrán vastagsága körülbelül 7,5 nm.
Membrán funkciók
A membránok a következő funkciókat látják el:
- a sejttartalom elkülönítése a külső környezettől,
- a sejt és a környezet közötti anyagcsere szabályozása,
- a sejt felosztása rekeszekre („rekeszek”),
- az „enzimatikus szállítószalagok” lokalizációjának helye,
- a sejtek közötti kommunikáció biztosítása a többsejtű szervezetek szöveteiben (adhézió),
- jelfelismerés.
A legfontosabb membrán tulajdonság— szelektív permeabilitás, i.e. a membránok bizonyos anyagok vagy molekulák számára nagyon áteresztők, mások számára pedig rosszul (vagy teljesen át nem eresztők) áteresztők. Ez a tulajdonság alapozza meg a membránok szabályozó funkcióját, biztosítva az anyagcserét a sejt és a külső környezet között. A sejtmembránon áthaladó anyagok folyamatát ún anyagok szállítása. Vannak: 1) passzív szállítás- az anyagok energiafogyasztás nélküli átadásának folyamata; 2) aktív közlekedés- az anyagok áthaladásának folyamata, amely energiafelhasználással történik.
at passzív szállítás az anyagok nagyobb koncentrációjú területről alacsonyabb koncentrációjú területre mozognak, pl. koncentráció gradiens mentén. Minden oldatban vannak oldószer és oldott anyag molekulák. Az oldott anyag molekulák mozgásának folyamatát diffúziónak, az oldószermolekulák mozgását ozmózisnak nevezzük. Ha a molekula feltöltött, akkor a transzportját is befolyásolja az elektromos gradiens. Ezért az emberek gyakran elektrokémiai gradiensről beszélnek, amely a két gradienst kombinálja. A szállítás sebessége a gradiens nagyságától függ.
A passzív szállításnak a következő típusai különböztethetők meg: 1) egyszerű diffúzió— anyagok szállítása közvetlenül a lipid kettős rétegen keresztül (oxigén, szén-dioxid); 2) diffúzió a membráncsatornákon keresztül— transzport csatornaképző fehérjéken (Na +, K +, Ca 2+, Cl -) keresztül; 3) megkönnyített diffúzió- anyagok szállítása speciális transzportfehérjék segítségével, amelyek mindegyike felelős bizonyos molekulák vagy rokon molekulacsoportok (glükóz, aminosavak, nukleotidok) mozgásáért; 4) ozmózis— vízmolekulák szállítása (minden biológiai rendszerben az oldószer a víz).
Szükség aktív közlekedés akkor fordul elő, amikor biztosítani kell a molekulák transzportját a membránon az elektrokémiai gradiens ellen. Ezt a transzportot speciális hordozófehérjék végzik, amelyek aktivitása energiafelhasználást igényel. Az energiaforrás az ATP molekulák. Az aktív transzport a következőket tartalmazza: 1) Na + /K + pumpa (nátrium-kálium pumpa), 2) endocitózis, 3) exocitózis.

Na + /K + szivattyú működése. A normális működéshez a sejtnek fenn kell tartania a K + és Na + ionok bizonyos arányát a citoplazmában és a külső környezetben. A sejten belüli K + koncentrációnak lényegesen magasabbnak kell lennie, mint azon kívül, és a Na + - fordítva. Megjegyzendő, hogy a Na + és a K + szabadon diffundálhat a membrán pórusain. A Na + /K + pumpa ellensúlyozza ezen ionok koncentrációjának kiegyenlítését, és aktívan pumpálja a Na +-t a sejtből, a K +-t pedig a sejtbe. A Na + /K + pumpa egy transzmembrán fehérje, amely képes konformációs változásokra, aminek következtében K + és Na + is képes megkötni. A Na + /K + pumpaciklus a következő fázisokra osztható: 1) Na + hozzáadása a membrán belsejéből, 2) a pumpa fehérje foszforilációja, 3) Na + felszabadulása az extracelluláris térben, 4) K + hozzáadása a membránon kívülről , 5) a pumpa fehérje defoszforilációja, 6) K + felszabadulása az intracelluláris térben. A sejtek működéséhez szükséges energia csaknem egyharmadát a nátrium-kálium pumpa működésére fordítják. Egy működési ciklusban a szivattyú 3Na +-t pumpál ki a cellából és 2K +-t pumpál be.
Endocitózis- a nagy részecskék és makromolekulák sejt általi felszívódásának folyamata. Az endocitózisnak két típusa van: 1) fagocitózis- nagy részecskék (sejtek, sejtrészek, makromolekulák) befogása és abszorpciója és 2) pinocytosis— folyékony anyagok (oldat, kolloid oldat, szuszpenzió) felfogása és felszívása. A fagocitózis jelenségét I.I. Mechnikov 1882-ben. Az endocitózis során a plazmamembrán invaginációt képez, élei összeolvadnak, és a citoplazmától egyetlen membránnal elhatárolt struktúrák befűződnek a citoplazmába. Számos protozoa és néhány leukocita képes fagocitózisra. Pinocitózis figyelhető meg a bélhámsejtekben és a vérkapillárisok endotéliumában.
Exocitózis- az endocitózissal ellentétes folyamat: különböző anyagok eltávolítása a sejtből. Az exocitózis során a hólyagmembrán egyesül a külső citoplazmatikus membránnal, a vezikula tartalma a sejten kívülre kerül, membránja pedig a külső citoplazmatikus membránba kerül. Ily módon a hormonok eltávolításra kerülnek a belső elválasztású mirigyek sejtjéből a protozoonokban, eltávolítják az emésztetlen táplálékmaradványokat.
Menj ide előadások 5. sz"Sejtelmélet. A sejtes szervezet típusai"
Menj ide előadások 7. sz"Eukarióta sejt: az organellumok szerkezete és funkciói"