기공이란 무엇이고 어디에 있나요? 기공의 보호세포와 이차세포
표피 조직 시스템에 속하는 기공은 식물의 생명에 특히 중요합니다. 기공의 구조는 매우 독특하고 그 중요성이 너무 커서 별도로 고려해야 합니다.
표피 조직의 생리학적 중요성은 이중적이며 대체로 모순적입니다. 한편으로, 표피는 구조적으로 식물이 건조되는 것을 방지하도록 적응되어 있으며, 이는 표피 세포의 단단한 폐쇄, 큐티클의 형성 및 상대적으로 긴 덮개 털에 의해 촉진됩니다. 그러나 반면에 표피는 서로 반대 방향으로 돌진하는 수증기와 다양한 가스 덩어리를 통과해야 합니다. 어떤 상황에서는 가스와 증기 교환이 매우 격렬해질 수 있습니다. 식물 유기체에서 이러한 모순은 기공의 도움으로 성공적으로 해결됩니다. 기공은 특이하게 변형된 두 개의 표피 세포로 구성되며, 기공은 반대쪽(길이를 따라) 끝으로 서로 연결되어 있으며 기공이라고 합니다. 보호 세포. 그들 사이의 세포 간 공간을 기공 균열.
가드 세포는 팽압의 주기적 변화를 통해 기공 틈이 열리거나 닫히는 방식으로 모양이 바뀌기 때문에 그렇게 불립니다. 다음 두 가지 특징은 이러한 기공 운동에 매우 중요합니다. 첫째, 공변세포는 표피의 다른 세포와 달리 엽록체를 함유하고 있는데, 엽록체에서 빛을 받으면 광합성이 일어나고 당이 형성된다. 삼투압 활성 물질인 당의 축적은 표피의 다른 세포에 비해 공변 세포의 팽압에 변화를 일으킵니다. 둘째, 공변 세포막은 고르지 않게 두꺼워지기 때문에 팽압의 변화로 인해 이들 세포의 부피가 고르지 않게 변하고 결과적으로 모양도 변합니다. 공변 세포의 모양 변화는 기공 틈의 폭 변화를 유발합니다. 다음 예를 통해 이를 설명해 보겠습니다. 그림은 쌍떡잎 식물의 기공 유형 중 하나를 보여줍니다. 기공의 가장 바깥쪽 부분은 큐티클에 의해 형성된 막질 돌기로 구성되며 때로는 중요하지 않으며 때로는 상당히 중요합니다. 그들은 외부 표면에서 작은 공간을 제한하며, 그 아래쪽 경계는 기공 간격 자체입니다. 앞마당 기공. 기공 틈 뒤에는 내부에 공변 세포 측벽의 작은 내부 돌기로 구분되는 또 다른 작은 공간이 있습니다. 안뜰 기공. 안뜰은 다음과 같은 큰 세포 간 공간으로 직접 열립니다. 공기 구멍.
빛 속에서 당은 공변 세포에서 형성되고, 인접한 세포에서 물을 끌어당기고 공변 세포의 팽압이 증가하며 껍질의 얇은 부분이 두꺼운 부분보다 더 늘어납니다. 따라서 기공 틈으로 돌출된 볼록한 돌기가 편평해지고 기공이 열립니다. 예를 들어 설탕이 밤에 전분으로 변하면 공변 세포의 팽압이 떨어지고 이로 인해 껍질의 얇은 부분이 약화되고 서로를 향해 튀어 나와 기공이 닫힙니다. 다른 식물에서는 기공 간격을 닫고 여는 메커니즘이 다를 수 있습니다. 예를 들어, 풀과 사초의 공변 세포는 끝이 넓어지고 중간 부분이 좁아집니다. 세포 중간 부분의 막은 두꺼워지고 확장된 끝 부분은 얇은 셀룰로오스 막을 유지합니다. 팽압이 증가하면 세포 끝이 부풀어 오르고 결과적으로 직선 중앙 부분이 서로 멀어집니다. 이로 인해 기공이 열리게 됩니다.
기공 장치의 작동 메커니즘의 특징은 공변 세포의 모양과 구조, 그리고 기공에 인접한 표피 세포의 참여에 의해 생성됩니다. 기공에 바로 인접한 세포가 표피의 다른 세포와 모양이 다른 경우 이를 기공이라고 합니다. 기공의 동반세포.
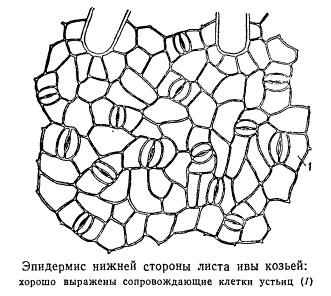
대부분의 경우 동반 셀과 후행 셀은 공통 기원을 갖습니다.
기공의 보호 세포는 표피 표면 위로 약간 올라가거나 반대로 다소 깊은 구덩이로 낮아집니다. 표피 표면의 일반적인 수준과 관련하여 공변 세포의 위치에 따라 기공 균열의 너비를 조정하는 메커니즘 자체가 다소 변경됩니다. 때때로 장루의 공변세포가 목화화되고, 기공열구의 개방 조절은 인접한 표피세포의 활동에 의해 결정됩니다. 팽창 및 수축, 즉 부피를 변경하여 인접한 공변 세포를 동반합니다. 그러나 종종 목질화된 공변 세포가 있는 기공은 전혀 닫히지 않습니다. 이러한 경우 가스 및 증기 교환 강도의 조절은 소위 건조 시작을 통해 다르게 수행됩니다. 목화 공변 세포가 있는 기공에서 큐티클은 종종 전체 기공 틈뿐만 아니라 공기 구멍까지 확장되어 바닥을 감싸는 상당히 두꺼운 층으로 덮이는 경우가 많습니다.
대부분의 식물은 잎의 양쪽에 기공이 있거나 밑면에만 기공이 있습니다. 그러나 잎의 윗면(물 표면에 떠 있는 잎)에만 기공이 형성되는 식물도 있습니다. 일반적으로 녹색 줄기보다 잎에 기공이 더 많습니다.
다양한 식물의 잎에 있는 기공 수는 매우 다양합니다. 예를 들어, 차양이 없는 브롬 잎 밑면의 기공 수는 1mm 2 당 평균 30개이며, 동일한 조건에서 자라는 해바라기에서는 약 250개입니다. 일부 식물은 1mm 2 당 최대 1300개의 기공을 가지고 있습니다.
동일한 식물 종의 표본에서 기공의 밀도와 크기는 환경 조건에 따라 크게 달라집니다. 예를 들어, 밝은 곳에서 자란 해바라기 잎에는 잎 표면 1mm 2 당 평균 220개의 기공이 있었고, 첫 번째 옆에 자란 표본에는 약간의 음영이 있었지만 약 140개가 있었습니다. 한 식물이 완전한 빛에서 자라면 기공의 밀도는 아래쪽 잎에서 위쪽 잎으로 증가합니다.
기공의 수와 크기는 식물의 성장 조건뿐만 아니라 식물 자체의 생명 과정의 내부 관계에 따라 크게 달라집니다. 이 값(계수)은 식물의 성장을 결정하는 각 요소 조합에 대해 가장 민감한 시약입니다. 따라서 다양한 조건에서 자란 식물 잎의 기공 밀도와 크기를 결정하면 각 식물과 환경의 관계 특성에 대한 아이디어를 얻을 수 있습니다. 특정 기관의 해부학적 요소의 크기와 수를 결정하는 모든 방법은 재배 식물의 각 품종이 단위 면적당 해부학적 요소의 크기와 수에 대한 특정 제한이 특징입니다. 정량적 해부학 방법은 식물 재배와 생태학 모두에서 큰 이점을 가지고 사용될 수 있습니다.
가스 및 증기 교환을 위한 기공과 함께 물이 증기 형태가 아닌 방울 액체 상태로 방출되는 기공도 있습니다. 때때로 그러한 기공은 일반 기공과 매우 유사하며 약간 더 크며 공변 세포에는 이동성이 부족합니다. 완전히 성숙한 상태의 기공에는 공변 세포가 없고 물을 밖으로 내보내는 구멍만 남는 경우가 많습니다. 액체 물방울을 분비하는 기공을 기공이라고 합니다. 물, 그리고 물방울-액체 물의 방출과 관련된 모든 형성 - 수소화물.
hydathodes의 구조는 다양합니다. 일부 포충류에는 물을 제거하는 구멍 아래에 실질조직이 있는데, 이는 물 전도 시스템에서 물을 이동하고 기관에서 방출하는 데 관여합니다. 다른 수돗물에서는 물 전도 시스템이 출구에 직접 접근합니다. 포충류는 특히 다양한 식물 묘목의 첫 번째 잎에 형성되는 경우가 많습니다. 따라서 습하고 따뜻한 날씨에는 곡물, 완두콩 및 많은 초원 풀의 어린 잎이 물방울을 한 방울 씩 방출합니다. 이 현상은 여름 전반기, 날씨가 좋은 날 이른 아침에 관찰할 수 있습니다.
가장 잘 정의된 포낭은 잎의 가장자리를 따라 위치합니다. 종종 하나 이상의 포식체가 잎의 가장자리를 끄는 각 치아에 의해 운반됩니다.
오류를 발견하면 텍스트 부분을 강조 표시하고 다음을 클릭하세요. Ctrl+Enter.
식물의 기공은 표피층에 위치한 모공입니다. 그들은 꽃과 환경 사이의 과도한 물과 가스 교환을 증발시키는 역할을 합니다.
그들은 1675년 박물학자 Marcello Malpighi가 자신의 발견을 Anatome plantarum에 발표하면서 처음으로 알려졌습니다. 그러나 그는 그들의 실제 목적을 밝히지 못했고, 이는 더 많은 가설과 연구 개발의 원동력이 되었습니다.
연구의 역사
19세기에는 오랫동안 기다려온 연구가 진전되었습니다. Hugo von Mohl과 Simon Schwendener 덕분에 기공 작동의 기본 원리와 구조 유형에 따른 분류가 알려졌습니다.
이러한 발견은 모공의 기능을 이해하는 데 강력한 자극을 주었지만 이전 연구의 일부 측면은 오늘날까지 계속해서 연구되고 있습니다.
잎의 구조
표피나 기공과 같은 식물 부분은 잎의 내부 구조와 관련이 있지만 먼저 잎의 외부 구조를 연구해야 합니다. 따라서 시트는 다음으로 구성됩니다.
- 잎사귀 - 광합성, 가스 교환, 수분 증발 및 식물 번식(특정 종의 경우)을 담당하는 평평하고 유연한 부분입니다.
- 성장판과 잎자루가 위치한 기부입니다. 또한 잎을 줄기에 부착하는 데 도움이 됩니다.
- 턱잎은 겨드랑이 싹을 보호하는 기저부의 한 쌍의 형성입니다.
- 잎자루 - 잎과 줄기를 연결하는 잎의 점점 가늘어지는 부분. 그것은 중요한 기능, 즉 교육 조직을 통한 빛과 성장을 향한 방향을 담당합니다.
잎의 외부구조는 모양과 종류(단순/복잡)에 따라 조금씩 다를 수 있으나, 위의 부분은 모두 항상 존재한다.
내부 구조에는 표피와 기공뿐만 아니라 다양한 형성 조직과 정맥이 포함됩니다. 각 요소에는 고유한 디자인이 있습니다.
예를 들어, 잎의 바깥쪽은 크기와 모양이 다른 살아있는 세포로 구성되어 있습니다. 그 중 가장 표면은 투명하여 햇빛이 잎에 침투할 수 있습니다.
약간 더 깊은 곳에 위치한 작은 세포에는 잎에 녹색을 주는 엽록체가 포함되어 있습니다. 그 속성으로 인해 폐쇄라고 불렸습니다. 수분의 정도에 따라 수축되거나 서로 사이에 기공 틈이 형성됩니다.

구조
식물의 기공 길이는 받는 빛의 종류와 정도에 따라 달라집니다. 가장 큰 모공 크기는 1cm에 달하며, 기공은 구멍의 수준을 조절하는 보호 세포를 형성합니다.
그들의 움직임 메커니즘은 매우 복잡하며 식물 종에 따라 다릅니다. 대부분의 경우 물 공급과 엽록체 수준에 따라 세포 조직의 팽압이 감소하거나 증가하여 기공 개방을 조절할 수 있습니다.

기공 균열의 목적
시트의 기능과 같은 측면에 대해 자세히 설명할 필요는 없을 것입니다. 남학생조차도 이것에 대해 알고 있습니다. 그러면 기공은 무엇을 담당합니까? 그들의 임무는 공변 세포의 작업을 통해 달성되는 증산(식물을 통한 물 이동 과정 및 잎, 줄기, 꽃과 같은 외부 기관을 통한 증발 과정)을 보장하는 것입니다. 이 메커니즘은 더운 날씨에 식물이 건조되는 것을 방지하고 과도한 습도 조건에서 부패 과정이 시작되는 것을 방지합니다. 작동 원리는 매우 간단합니다. 세포의 체액량이 충분히 높지 않으면 벽에 가해지는 압력이 떨어지고 기공 틈이 닫혀 생명을 유지하는 데 필요한 수분 함량이 유지됩니다.
반대로, 그 초과분은 압력을 증가시키고 과도한 수분이 증발하는 모공을 열어줍니다. 이로 인해 냉각 시설에서 기공의 역할도 큽니다. 왜냐하면 기공 주변의 공기 온도가 증산을 통해 정확하게 감소하기 때문입니다.
또한 틈새 아래에는 가스 교환에 사용되는 공기 구멍이 있습니다. 공기는 모공을 통해 식물 안으로 들어가 호흡을 시작합니다. 그러면 과잉 산소는 동일한 기공 틈을 통해 대기 중으로 빠져나갑니다. 또한 식물을 분류하는 데에는 그 유무를 사용하는 경우가 많습니다.

워크시트 기능
잎은 광합성, 호흡, 증산, 소화, 영양 번식을 수행하는 외부 기관입니다. 또한 기공을 통해 수분과 유기물을 축적할 수 있으며, 식물이 어려운 환경 조건에 더 잘 적응할 수 있도록 해줍니다.
물은 주요 세포내 매체이기 때문에 나무나 꽃 내부의 체액 배설과 순환은 나무나 꽃의 생명에 똑같이 중요합니다. 이 경우 식물은 통과하는 모든 수분의 0.2 %만을 흡수하고 나머지는 증산 및 소화로 이동하여 용해 된 미네랄 염의 이동 및 냉각이 발생합니다.
영양번식은 꽃잎을 자르고 뿌리를 내리면서 일어나는 경우가 많습니다. 많은 실내 식물이 이런 방식으로 재배됩니다. 이것이 품종의 순도를 유지하는 유일한 방법이기 때문입니다.
앞서 언급했듯이 다양한 자연 조건에 적응하는 데 도움이 됩니다. 예를 들어, 가시로의 변형은 사막 식물이 수분 증발을 줄이는 데 도움이 되고, 덩굴손은 줄기의 기능을 강화하며, 큰 크기는 종종 기후 조건으로 인해 정기적으로 보충이 불가능한 액체와 영양분을 보존하는 역할을 합니다.
그리고 이 목록은 끝없이 계속될 수 있습니다. 동시에 이러한 기능이 꽃과 나무의 잎에서도 동일하다는 사실을 눈치 채지 못하는 것은 어렵습니다.

기공이 없는 식물은 무엇입니까?
기공 틈은 고등 식물의 특징이므로 모든 종에 존재하며, 나무나 꽃에 잎이 없더라도 없다고 생각하는 것은 실수입니다. 규칙의 유일한 예외는 다시마와 기타 조류입니다.
기공의 구조와 침엽수, 양치류, 속새, 수영식물에서의 작용은 꽃 피는 식물의 구조와 다릅니다. 대부분의 경우 낮에는 슬릿이 열려 있어 가스 교환 및 증산에 적극적으로 참여합니다. 건조한 지역에서 수분을 보존하기 위해 밤에는 모공이 열리고 아침에는 닫히는 선인장과 다육식물은 예외입니다.
잎이 물 표면에 떠있는 식물의 기공은 표피의 상층에만 위치하고 "고착성"잎 - 하층에만 위치합니다. 다른 종류에서는 이러한 슬롯이 플레이트의 양쪽에 있습니다.
기공 위치
기공 틈새는 잎사귀의 양쪽에 있지만 아래쪽 부분의 수가 위쪽 부분보다 약간 더 많습니다. 이러한 차이는 조명이 밝은 시트 표면에서 수분 증발을 줄여야 하기 때문에 발생합니다.
단자엽 식물의 경우 기공 위치는 판의 성장 방향에 따라 달라지므로 특이성이 없습니다. 예를 들어, 수직 방향 식물 잎의 표피에는 상층과 하층 모두에 동일한 수의 기공이 있습니다.

앞서 언급했듯이 떠 다니는 잎은 밑면에 기공 틈이 없습니다. 왜냐하면 기공이 전혀없는 완전 수생 식물처럼 큐티클을 통해 수분을 흡수하기 때문입니다.
침엽수의 기공은 내피 아래 깊숙이 위치하여 증산 능력의 감소에 기여합니다.
또한 모공의 위치는 표피 표면에 따라 다릅니다. 틈은 나머지 "피부" 세포와 수평을 이루거나, 더 높거나 더 낮거나, 규칙적인 줄을 형성하거나, 외피 조직 전체에 무작위로 흩어져 있을 수 있습니다.
선인장, 다육 식물 및 잎이 없거나 변화하여 바늘 모양으로 변하는 기타 식물의 경우 기공이 줄기와 다육 부분에 있습니다.
유형
식물의 기공은 동반 세포의 위치에 따라 여러 유형으로 나뉩니다.
- Anomocytic - 부산물이 표피에서 발견되는 다른 부산물과 다르지 않은 가장 일반적인 것으로 간주됩니다. 간단한 변형 중 하나는 laterocyte 유형이라고 할 수 있습니다.
- Paracytic - 기공 틈에 대해 동반 세포가 평행하게 지대주하는 것이 특징입니다.
- Diacitic - 측면 입자가 두 개만 있습니다.
- 이형성(Anisocytic) - 꽃 피는 식물에서만 발견되는 유형으로, 3개의 동반 세포가 있으며, 그 중 하나는 크기가 눈에 띄게 다릅니다.
- 사핵세포(Tetracytic) - 외떡잎 식물의 특징이며 4개의 동반 세포가 있습니다.
- 백혈구 - 측면 입자가 닫히는 입자 주위의 고리로 닫힙니다.
- Pericytic - 동반 세포에 연결되지 않은 기공이 특징입니다.
- Desmocyte - 간격과 측면 입자 사이에 접착이 있는 경우에만 이전 유형과 다릅니다.
여기에는 가장 인기 있는 유형만 나열되어 있습니다.
잎의 외부 구조에 대한 환경 요인의 영향
식물의 생존을 위해서는 적응성의 정도가 매우 중요합니다. 예를 들어, 습한 지역은 잎이 크고 기공이 많은 것이 특징인 반면, 건조한 지역에서는 이 메커니즘이 다르게 작동합니다. 꽃이나 나무 모두 크기가 다르지 않고, 모공의 수가 눈에 띄게 줄어들어 과도한 증발을 막아줍니다.

따라서 환경의 영향으로 식물의 일부가 시간이 지남에 따라 어떻게 변화하는지 추적할 수 있으며, 이는 기공 수에도 영향을 미칩니다.
실내 식물의 기공 상태 확인
식물의 잎은 다양한 기능을 수행합니다. 이는 광합성, 가스 교환 및 증산(물 증발)이 일어나는 주요 기관입니다. 가스 교환을 수행하기 위해 식물의 육상 기관에는 기공이라는 특별한 구조가 있습니다.
기공은 표피(잎 피부)의 일부이지만 특별한 세포 그룹입니다. 기공 장치는 두 개의 보호 세포로 구성되며, 그 사이에는 기공 틈, 2-4개의 기공 주위 세포 및 기공 틈 아래에 위치한 가스-공기 챔버가 있습니다.
기공의 보호 세포는 길쭉하고 구부러진 "콩 모양" 모양을 가지고 있습니다. 기공 틈을 향한 벽이 두꺼워졌습니다. 기공 세포는 모양을 변경할 수 있습니다. 이로 인해 기공 균열이 열리거나 닫힙니다. 이 세포에는 엽록체(녹색 색소체)가 포함되어 있습니다. 기공 틈의 개폐는 공변 세포의 팽압(삼투압) 변화로 인해 발생합니다. 공변 세포의 엽록체에는 설탕으로 전환될 수 있는 전분이 포함되어 있습니다. 전분이 설탕으로 전환되면 삼투압이 증가하고 기공이 열립니다. 당 함량이 감소하면 역과정이 일어나 기공이 닫힙니다.
기공 틈은 종종 이른 아침에 활짝 열렸다가 낮 동안에는 닫혀 있습니다(또는 반 닫혀 있습니다). 기공 수는 환경 조건(온도, 빛, 습도)에 따라 다릅니다. 하루 중 다른 시간에 열리는 정도는 종에 따라 크게 다릅니다. 습한 서식지에 있는 식물의 잎에서 기공의 밀도는 1mm2당 100~700개입니다.
대부분의 육상 식물에서 기공은 잎의 밑면에서만 발견됩니다. 예를 들어 양배추나 해바라기처럼 잎의 양쪽에 있을 수도 있습니다. 또한 잎 위쪽과 아래쪽의 기공 밀도는 동일하지 않습니다. 양배추의 경우 1mm2 당 140과 240이고 해바라기의 경우 1mm2 당 175와 325입니다. 수련과 같은 수생식물의 기공은 잎의 윗면에만 위치하며 밀도는 1mm2당 약 500개이다. 수중 식물에는 기공이 전혀 없습니다.
작업의 목표:
다양한 실내 식물의 기공 상태 확인.
작업
1. 추가 문헌을 사용하여 다양한 식물의 기공 구조, 위치 및 수에 대한 질문을 연구합니다.
2. 연구할 식물을 선택하세요.
3. 생물학 교실에서 사용할 수 있는 다양한 실내 식물에서 기공의 상태와 개방 정도를 결정합니다.
재료 및 방법
기공 상태의 판정은 "식물 생리학에 대한 방법론적 권고사항"(E.F. Kim 및 E.N. Grishina 편찬)에 기술된 방법에 따라 수행하였다. 이 기술의 핵심은 기공의 개방 정도가 특정 화학 물질이 잎의 과육에 침투하여 결정된다는 것입니다. 이를 위해 에테르, 알코올, 가솔린, 등유, 벤젠, 자일렌 등 다양한 액체가 사용됩니다. 화학실에서 제공되는 알코올, 벤젠, 자일렌을 사용했습니다. 이러한 액체가 잎 속으로 침투하는 정도는 기공이 열리는 정도에 따라 달라집니다. 잎 뒷면에 액체 한 방울을 떨어뜨린 후 2~3분 후에 잎에 밝은 반점이 나타나면 액체가 기공을 통과했다는 의미입니다. 이 경우 알코올은 넓게 열린 기공, 벤젠을 통해서만 잎에 침투합니다. 이미 평균 개구부 너비가 있고 자일렌 만 거의 닫힌 기공을 통해 침투합니다.
작업의 첫 번째 단계에서 우리는 다양한 식물의 기공 상태(개방 정도)를 결정할 수 있는 가능성을 확립하려고 노력했습니다. 이 실험에서는 Agave, cyperus, tradescantia, geranium, oxalis, syngonium, Amazonian lily, begonia, sanchetia, dieffenbachia, clerodendron, passionflower, 호박 및 콩을 사용했습니다. 추가 작업을 위해 옥살리스, 제라늄, 베고니아, 산케티아, 클레로덴드론, 시계꽃, 호박 및 콩을 선택했습니다. 다른 경우에는 기공 개방 정도를 확인할 수 없었습니다. 이는 용설란, 사이페러스, 백합의 잎이 기공 틈새를 통해 물질이 침투하는 것을 방지하는 코팅으로 덮여 있기 때문일 수 있습니다. 또 다른 가능한 이유는 실험 시간(14시)에 기공이 이미 닫혀 있었기 때문일 수 있습니다.
연구는 일주일에 걸쳐 진행됐다. 매일 방과 후 14시에 위의 방법을 이용하여 기공 개방 정도를 측정하였다.
결과 및 토론
얻은 데이터는 표에 나와 있습니다. 주어진 데이터는 평균화되었기 때문에 날마다 기공의 상태가 달랐습니다. 따라서 6번의 측정 결과, 기공이 넓게 열리는 정도는 나무밤색에서는 2번, 제라늄에서는 1번, 베고니아에서는 평균 기공 개방 정도가 2번 기록되었다. 이러한 차이는 실험 시간에 따라 달라지지 않습니다. 아마도 사무실의 온도와 식물의 조명은 상당히 일정했지만 기후 조건과 관련이 있을 것입니다. 따라서 얻은 평균 데이터는 이러한 식물에 대한 특정 표준으로 간주될 수 있습니다.
이 연구는 동시에, 동일한 조건 하에서 서로 다른 식물에서 기공 개방 정도가 동일하지 않음을 나타냅니다. 기공이 넓게 열려 있는 식물(베고니아, 산체티아, 호박)과 중간 크기의 기공 틈(옥살리스, 제라늄, 콩)이 있는 식물이 있습니다. 좁은 기공 틈새는 클레로덴드론에서만 발견됩니다.
우리는 이러한 결과를 예비적인 것으로 간주합니다. 앞으로는 다양한 식물의 기공 개폐에 생물학적 리듬이 존재하는지, 어떻게 존재하는지 규명할 계획이다. 이를 위해 낮에는 기공 균열 상태를 모니터링합니다.
Stomata, 그 구조 및 작용 메커니즘
표피 세포는 외벽의 독특한 구조로 인해 물과 가스가 거의 침투할 수 없습니다. 플랜트와 외부 환경 간의 가스 교환 및 물 증발은 어떻게 수행됩니까? 플랜트의 정상적인 기능에 필요한 프로세스는 무엇입니까? 표피 세포 중에는 기공이라고 불리는 특징적인 형성이 있습니다.
기공은 틈새 모양의 개구부로, 대부분 초승달 모양인 두 개의 보호 세포로 양쪽 경계를 이루고 있습니다.
기공은 가스 교환이 일어나는 표피의 구멍입니다. 주로 잎에서 발견되지만 줄기에서도 발견됩니다. 각 기공은 다른 표피 세포와 달리 엽록체를 포함하는 보호 세포로 양쪽이 둘러싸여 있습니다. 공변 세포는 부풀어오르는 정도를 변경하여 기공 개구부의 크기를 조절합니다.
이 세포는 살아 있으며 표피의 다른 세포에는 없는 엽록소 알갱이와 전분 알갱이를 포함하고 있습니다. 특히 잎에 기공이 많이 있다. 단면은 잎 조직 내부의 기공 바로 아래에 호흡강이라고 불리는 구멍이 있음을 보여줍니다. 그 틈 내에서 가드셀은 가운데 부분에서 서로 가까워지고, 위쪽과 아래쪽은 더 떨어져서 앞마당과 뒷마당이라는 공간을 형성합니다.
공변 세포는 크기를 늘리거나 줄일 수 있기 때문에 기공 틈이 때로는 넓게 열리기도 하고, 때로는 좁아지거나 심지어 완전히 닫히기도 합니다.
따라서 공변 세포는 기공을 열고 닫는 과정을 조절하는 장치입니다.
이 과정은 어떻게 수행되나요?
틈새를 향한 공변 세포의 벽은 인접한 표피 세포를 향한 벽보다 훨씬 두껍습니다. 식물에 빛을 비추고 수분이 과잉되면 전분은 공변 세포의 엽록소 알갱이에 축적되며, 그 중 일부는 설탕으로 전환됩니다. 세포 수액에 용해된 설탕은 인접한 표피 세포로부터 물을 끌어당기며, 그 결과 공변 세포의 팽압이 증가합니다. 강한 압력은 표피에 인접한 세포벽의 돌출을 가져오고, 반대쪽의 크게 두꺼워진 벽은 곧게 펴집니다. 결과적으로 기공 틈이 열리고 가스 교환과 수분 증발이 증가합니다. 어둡거나 습기가 부족한 경우 팽압이 감소하고 공변 세포가 이전 위치로 돌아가고 두꺼운 벽이 닫힙니다. 기공 틈이 닫힙니다.
기공은 식물의 모든 젊고 발화되지 않은 지상 기관에 위치합니다. 특히 잎에 많이 있으며 여기서는 주로 아래쪽 표면에 위치합니다. 잎이 수직으로 놓여지면 양쪽에 기공이 발달합니다. 물 표면에 떠 있는 일부 수생 식물(예: 수련, 난낭)의 잎에서는 기공이 잎의 윗면에만 위치합니다.
1제곱미터당 기공 수 잎 표면의 mm는 평균 300mm이지만 때로는 600 이상에 이릅니다. 부들(Typha)은 1평방미터당 1300개 이상의 기공을 가지고 있습니다. mm. 물에 잠긴 잎에는 기공이 없습니다. 기공은 피부 전체 표면에 고르게 위치하는 경우가 가장 많지만 일부 식물에서는 그룹으로 수집됩니다. 단자엽 식물과 많은 침엽수의 바늘에 세로줄로 위치합니다. 건조한 지역의 식물에서는 기공이 잎 조직에 잠겨 있는 경우가 많습니다. 기공 발달은 일반적으로 다음과 같이 발생합니다. 표피의 개별 세포에는 아치형 벽이 형성되어 세포를 여러 개의 작은 세포로 나누어 중앙 세포가 기공의 조상이 됩니다. 이 세포는 세로(세포 축을 따라) 격막으로 나누어집니다. 이 격막은 갈라지고 틈이 형성됩니다. 이를 제한하는 세포는 기공의 보호 세포가 됩니다. 일부 간 이끼에는 공변 세포가 없는 독특한 기공이 있습니다.
그림에서. 주사전자현미경을 이용하여 얻은 현미경 사진에서 기공과 공변세포의 모습을 보여준다.
여기서는 공변 세포의 세포벽 두께가 이질적이라는 것을 알 수 있습니다. 기공 입구에 더 가까운 벽은 반대쪽 벽보다 확실히 더 두껍습니다. 또한 세포벽을 구성하는 셀룰로오스 미세섬유는 구멍을 향한 벽이 덜 탄력적이도록 배열되어 있으며 일부 섬유는 소시지와 유사하게 공변 세포 주위에 일종의 고리를 형성합니다. 세포가 물을 흡수하여 부풀어 오르면 이 고리는 세포가 더 이상 팽창하는 것을 방지하고 길이만 늘어나게 합니다. 공변 세포는 끝 부분이 연결되어 있고 기공 틈에서 떨어진 얇은 벽이 더 쉽게 늘어나므로 세포는 반원형 모양을 얻습니다. 따라서 공변 세포 사이에 구멍이 나타납니다. (소시지 모양 풍선의 한쪽 면을 따라 접착 테이프를 붙인 채로 부풀려도 같은 효과를 얻을 수 있습니다.)
반대로, 물이 공변 세포를 떠나면 기공이 닫힙니다. 세포 팽창의 변화가 어떻게 발생하는지는 아직 명확하지 않습니다.
전통적인 가설 중 하나인 "설탕-전분" 가설은 낮 동안 공변 세포의 설탕 농도가 증가하고 결과적으로 세포의 삼투압과 세포 내로의 물 흐름이 증가한다고 제안합니다. 그러나 아직까지 공변 세포에 충분한 당이 축적되어 삼투압의 변화가 관찰된다는 사실을 보여준 사람은 아무도 없습니다. 최근에는 낮 동안 빛 속에서 칼륨 이온과 그에 수반되는 음이온이 공변 세포에 축적된다는 사실이 밝혀졌습니다. 이러한 이온 축적은 관찰된 변화를 일으키기에 충분합니다. 어둠 속에서 칼륨 이온(K+)은 보호 세포를 떠나 인접한 표피 세포로 이동합니다. 어떤 음이온이 칼륨 이온의 양전하와 균형을 이루는지는 아직 불분명합니다. 연구된 식물 중 일부(전부는 아님)는 말산염과 같은 유기산의 음이온이 다량 축적된 것으로 나타났습니다. 동시에, 공변 세포의 엽록체에서 어둠 속에서 나타나는 전분 입자의 크기가 감소합니다. 이는 전분이 빛에 노출되면 말산염으로 전환된다는 것을 의미합니다.
Allium cepa(양파)와 같은 일부 식물은 공변 세포에 전분이 없습니다. 따라서 기공이 열리면 말산염이 축적되지 않고, 염화물(Cl-)과 같은 무기 음이온과 함께 양이온이 흡수되는 것으로 보인다.
일부 질문은 해결되지 않은 상태로 남아 있습니다. 예를 들어, 기공을 열려면 왜 빛이 필요한가요? 엽록체는 전분을 저장하는 것 외에 어떤 역할을 합니까? 말산염은 어둠 속에서 다시 전분으로 변합니까? 1979년에 Vicia faba(파바콩)의 공변 세포의 엽록체에는 캘빈 회로 효소가 부족하고 엽록소는 존재하지만 틸라코이드 시스템은 제대로 발달되지 않은 것으로 나타났습니다. 결과적으로 광합성의 일반적인 C3 경로가 작동하지 않고 전분이 형성되지 않습니다. 이는 일반 광합성 세포처럼 낮에 전분이 형성되지 않고 밤에 형성되는 이유를 설명하는 데 도움이 될 수 있습니다. 또 다른 흥미로운 사실은 공변 세포에 플라스모데스마타(plasmodesmata)가 없다는 것입니다. 표피의 다른 세포로부터 이들 세포를 비교 분리합니다.
동정.
Stomata는 두 가지 주요 기능을 수행합니다. 가스 교환과 증산(증발)을 수행합니다.
기공은 두 개의 보호 세포와 그 사이에 있는 기공 틈으로 구성됩니다. 공변 세포 옆에는 2차(방구공) 세포가 있습니다. 기공 아래에는 공기 구멍이 있습니다. 기공은 필요에 따라 자동으로 닫히거나 열릴 수 있습니다. 이는 팽압 현상 때문입니다.
기공이 열리는 정도는 빛의 강도, 잎의 수분량, 이산화탄소의 양에 따라 달라집니다. 세포 간 공간, 기온 및 기타 요인. 운동 메커니즘을 유발하는 요인(잎 조직의 빛 또는 수분 결핍의 시작)에 따라 기공의 광 및 수력 운동이 구별됩니다. 또한 표피 세포의 수분 함량 변화로 인한 정수압 운동이 있으며 공변 세포의 대사에는 영향을 미치지 않습니다. 예를 들어, 깊은 물 부족으로 인해 잎이 시들고 표피 세포의 크기가 감소하여 보호 세포가 늘어나 기공이 열릴 수 있습니다. 또는 반대로 비가 내린 직후에는 표피 세포가 너무 많이 부풀어 오른다.
물이 공변 세포를 압축하고 기공이 닫힙니다.
하이드로패시브 솔루션 - 세포 실질이 물로 가득 차서 말단 세포를 기계적으로 압축할 때 기공 슬릿을 닫습니다.
수력적 개폐는 기공 공변 세포의 수분 함량 변화로 인해 발생하는 움직임입니다.
광활성 - 빛 속에서 기공이 열리고 어둠 속에서 닫히면서 나타납니다.
13. 증산에 대한 외부 요인의 영향
증산은 기공의 도움으로 수행되는 잎 표면이나 식물의 다른 부분에서 물이 증발하는 형태의 수분 손실입니다. 토양에 물이 부족하면 증산 속도가 감소합니다.
낮은 온도효소를 비활성화하여 물 흡수를 어렵게 만들고 증산 속도를 늦춥니다. 온도가 높으면 잎이 과열되어 증산량이 증가합니다. 온도가 증가함에 따라 증산 속도가 증가합니다. 온도는 물 증발을 위한 에너지원입니다. 증산에 의한 냉각 효과는 특히 고온, 낮은 공기 습도 및 양호한 물 공급에서 중요합니다. 또한 온도는 기공 개방 정도에 영향을 미치는 조절 기능도 수행합니다.
빛. 빛에서는 잎의 온도가 상승하고 증산량이 증가하며, 빛의 생리학적 효과는 기공의 움직임에 미치는 영향입니다. 식물은 어둠 속에서보다 빛 속에서 더 강하게 증산합니다. 증산에 대한 빛의 영향은 주로 녹색 세포가 적외선 햇빛뿐만 아니라 광합성에 필요한 가시광선도 흡수한다는 사실에 기인합니다. 완전한 어둠 속에서 기공은 처음에는 완전히 닫혔다가 약간 열립니다.
바람수증기의 동반으로 인해 증산이 증가하여 잎 표면에 결핍이 발생합니다. 풍속은 자유수면에서의 증발만큼 증산에 영향을 미치지 않습니다. 처음에 바람이 나타나고 속도가 증가하면 증산량이 증가하지만, 바람이 더 강해지면 이 과정에 거의 영향을 미치지 않습니다.
공기 습도. 과도한 습도로 인해 증산이 감소하고(온실에서) 건조한 공기에서는 증가합니다. 공기의 상대 습도가 낮을수록 수분 포텐셜이 낮아지고 증산이 더 빨라집니다. 잎에 물이 부족하면 기공 및 기공 외 조절이 발생합니다. 활성화되므로 증산의 강도는 수면에서 물이 증발하는 것보다 느리게 증가합니다. 심각한 물 부족이 발생하면 공기가 건조해짐에도 불구하고 증산이 거의 멈출 수 있습니다. 공기 습도가 증가하면 증산량이 감소합니다. 습도가 높으면 Guttation만 발생합니다.
높은 습도는 정상적인 증산 과정을 방해하므로 혈관을 통한 물질의 상향 이동, 식물 온도 조절 및 기공 운동에 부정적인 영향을 미칩니다.
물 결핍은 식물에 물이 부족한 것입니다.



