Kdo formuloval zákon zárodečné podobnosti. Vztah onto-fylogeneze
Zákon zárodečné podobnosti EMBRYOLOGIE ZVÍŘAT
Obecná embryologie: Terminologický slovník - Stavropol. O.V. Dileková, T.I. Lapina. 2010 .
Podívejte se, co je „zákon zárodečné podobnosti“ v jiných slovnících:
EMBRYOLOGIE ZVÍŘAT BAERŮV ZÁKON, ZÁKON ZÁRODKOVÉ PODOBNOSTI - zákon formuloval Carl Ernest von Baer v roce 1828: Společné základní rysy charakteristické pro každou velkou skupinu zvířat se objevují v procesu vývoje dříve než... ... Obecná embryologie: Terminologický slovník
Biogenetický zákon- Haeckel Muller (také známý jako „Haeckelův zákon“, „Haeckelův Mullerův zákon“, „Haeckelův zákon Darwina Mullera“, „základní biogenetický zákon“): každý živý tvor se ve svém individuálním vývoji (ontogenezi) opakuje ve známém ... . .. Wikipedie
BIOGENETICKÉ PRÁVO- (z řeckého bios - život a geneze - původ) stanovisko, které poprvé formulovali Fritz Müller a Ernst Haeckel, že řada forem, kterými prochází živá bytost v procesu svého individuálního vývoje od vajíčka k vyvinutému... ... Filosofická encyklopedie
Srovnávací anatomie zvířat- srovnávací morfologie, věda, která studuje zákonitosti stavby a vývoje orgánů a jejich systémů srovnáváním živočichů různých systematických skupin. Porovnání struktury orgánů v souvislosti s jejich funkcemi umožňuje pochopit... ... Velká sovětská encyklopedie
SROVNÁVACÍ ANATOMIE ZVÍŘAT- srovnávací morfologie, část morfologie živočichů, která studuje zákonitosti stavby a vývoje orgánů a jejich systémů srovnáváním živočichů různé systematiky. skupiny. Porovnání stavby orgánů v souvislosti s jejich funkcemi umožňuje... ... Biologický encyklopedický slovník
Důkazy evoluce- Fosilní archaeopteryx, objevený krátce po zveřejnění „... Wikipedie
Makroevoluce- organického světa je proces formování velkých systematických celků: z druhů nových rodů, z rodů nových čeledí atd. Makroevoluce je založena na stejných hnacích silách jako mikroevoluce: ... ... Wikipedia
embryologie- A; a. [z řečtiny embryonální embryo a doktrína logos] Obor biologie, který studuje tvorbu a vývoj embryí. E. zvířata. Srovnávací e. ◁ Embryologické, oh, oh. Tento výzkum. * * * embryologie (z embrya a...logie), nauka o... ... encyklopedický slovník
Evoluční doktrína- (také evolucionismus a evolucionismus) systém myšlenek a konceptů v biologii, které potvrzují historický progresivní vývoj biosféry Země, jejích biogeocenóz, jakož i jednotlivých taxonů a druhů, které lze zařadit do ... ... Wikipedia
Historie evolučního učení- má původ ve starověkých filozofických systémech, jejichž myšlenky byly zase zakořeněny v kosmologických mýtech. Impulsem k uznání evoluce vědeckou komunitou bylo vydání knihy Charlese Darwina „O původu druhů přirozenými prostředky... ... Wikipedia
Ontogeneze – implementace genetické informace ve všech fázích.
Ontogeneze je geneticky řízený proces. Během ontogeneze se realizuje genotyp a formuje se fenotyp.
Ontogeneze je individuální vývoj organismu, soubor postupných morfologických, fyziologických a biochemických přeměn, kterými organismus prochází od okamžiku svého vzniku až do konce života. O. zahrnuje růst, to znamená zvýšení tělesné hmotnosti, její velikost a diferenciaci. Výraz "O." zavedl E. Haeckel (1866), když formuloval biogenetický zákon.
První pokus o historické zdůvodnění O. učinil I. f. Meckel. Problém vztahu mezi O. a fylogenezí nastolil C. Darwin a rozvinuli F. Müller, E. Haeckel a další.Všechny nové evoluční znaky spojené se změnami dědičnosti vznikají u O., ale pouze ty, které přispívají k lepší adaptaci organismu na životní podmínky, jsou zachovány v procesu přirozeného výběru a přenášeny do dalších generací, tj. jsou fixní v evoluci. Znalost zákonitostí, příčin a faktorů kyslíku slouží jako vědecký základ pro hledání prostředků k ovlivnění vývoje rostlin, živočichů a člověka, což je nanejvýš důležité pro praxi rostlinného a živočišného původu, stejně jako pro lékařství. .
Fylogeneze je historický vývoj organismů. Termín byl zaveden v němčině. evolucionista E. Haeckel v roce 1866. Hlavním úkolem studia fyziologie je rekonstruovat evoluční přeměny živočichů, rostlin a mikroorganismů a na tomto základě stanovit jejich původ a související vztahy mezi taxony, ke kterým studované organismy patří. Za tímto účelem vyvinul E. Haeckel metodu „trojného paralelismu“, která umožňuje porovnáním dat tří věd – morfologie, embryologie a paleontologie – rekonstruovat průběh historického vývoje zkoumané systematické skupiny.
Zákon zárodečné podobnosti
Průzkumníci počátku 19. století. poprvé začali věnovat pozornost podobnosti fází vývoje embryí vyšších živočichů s fázemi složitosti organizace vedoucí od nízko organizovaných forem k progresivním. Při porovnání fází vývoje embryí různých druhů a tříd strunatců učinil K. Baer následující závěry.
1. Embrya zvířat stejného typu jsou v raných fázích vývoje podobná.
2. Důsledně se pohybují ve svém vývoji od obecnějších charakteristik typu ke stále specifičtějším. Nakonec se vyvinou znaky, které naznačují, že embryo patří k určitému rodu, druhu a nakonec k jednotlivým znakům.
3. Postupně se od sebe oddělují embrya různých zástupců stejného typu.
K. Baer, který nebyl evolucionistou, nedokázal spojit vzorce individuálního vývoje, které objevil, s procesem fylogeneze. Proto zobecnění, která učinil, měla význam pouze empirických pravidel.
Následný vývoj evoluční představy umožnil vysvětlit podobnost raných embryí jejich historickou příbuzností a jejich nabýváním stále specifičtějších rysů s postupnou vzájemnou izolací - skutečnou izolací odpovídajících tříd, řádů, čeledí, rody a druhy v procesu evoluce.
Charles Darwin brzy po objevu zákona embryonální podobnosti ukázal, že tento zákon svědčí o společném původu a jednotě počátečních fází evoluce v rámci typu.
Biogenetický zákon Haeckel-Müller: každý živý tvor ve svém individuálním vývoji (ontogenezi) do určité míry opakuje formy, které předali jeho předci nebo jeho druh (fylogeneze).
Ontogeneze - opakování fylogeneze
Srovnáním ontogeneze korýšů s morfologií jejich vyhynulých předků dospěl F. Muller k závěru, že žijící korýši ve svém vývoji opakují cestu, kterou urazili jejich předci. Transformace ontogeneze v evoluci se podle F. Mullera uskutečňuje díky jejímu prodloužení tím, že se k ní přidávají další stupně nebo extenze. Na základě těchto pozorování, jakož i studia vývoje strunatců, formuloval E. Haeckel (1866) základní biogenetický zákon, podle kterého Ontogeneze je krátké a rychlé opakování fylogeneze.
Opakování struktur charakteristických pro předky v embryogenezi potomků se nazývá rekapitulace. Rekapitulujte nejen morfologické znaky - notochorda, žaberní štěrbiny a žaberní oblouky u všech strunatců, ale i znaky biochemické organizace a fyziologie. Při evoluci obratlovců tedy dochází k postupné ztrátě enzymů nezbytných pro odbourávání kyseliny močové, produktu metabolismu purinů. U většiny bezobratlých je konečným produktem odbourávání kyseliny močové amoniak, u obojživelníků a ryb močovina, u mnoha plazů alantoin a u některých savců se kyselina močová vůbec nerozkládá a je vylučována močí. V embryogenezi savců a lidí jsou zaznamenány biochemické a fyziologické rekapitulace: uvolňování amoniaku ranými embryi, později močoviny, pak alantoinu a v posledních fázích vývoje - kyseliny močové.
V ontogenezi vysoce organizovaných organismů však není vždy dodržováno striktní opakování etap historického vývoje, jak vyplývá z biogenetického zákona. Lidské embryo tedy nikdy neopakuje dospělá stádia ryb, obojživelníků, plazů a savců, ale je v řadě znaků podobné pouze jejich embryím. Raná stádia vývoje zůstávají nejkonzervativnější, díky čemuž rekapitulují úplněji než ty pozdější. To je způsobeno skutečností, že jedním z nejdůležitějších mechanismů pro integraci raných fází embryogeneze je embryonální indukce a struktury embrya, které se tvoří jako první, jako je notochord, nervová trubice, hltan, střevo a somity, představují organizačních center embrya, od kterých závisí celý průběh vývoje.
Genetický základ rekapitulace spočívá v jednotě mechanismů genetického řízení vývoje, která je zachována na základě společných genů pro regulaci ontogeneze, které dědí příbuzné skupiny organismů po společných předcích.
Rekapitulace(z latinského recapitulatio - opakování) - pojem používaný v biologii k označení opakování v individuálním vývoji vlastností charakteristických pro dřívější fázi evolučního vývoje.
Lístek 96.
Ontogeneze jako základ fylogeneze. Cenogeneze. Autonomie ontogeneze. Filembryogeneze. Doktrína A. N. Severtsova o fylembryogenezi. Mechanismy jejich vzniku. Heterochronie a heterotopie biologických struktur ve vývoji ontogeneze.
Spoléháme-li se pouze na základní biogenetický zákon, je nemožné vysvětlit proces evoluce: nekonečné opakování toho, co bylo předáno, samo o sobě nezrodí něco nového. Jelikož život na Zemi existuje díky střídání generací konkrétních organismů, dochází k jeho evoluci díky změnám v jejich ontogenezích. Tyto změny se scvrkávaly na skutečnost, že specifické ontogeneze se odchylují od cesty vytyčené rodovými formami a získávají nové rysy.
Mezi takové odchylky patří např. cenogeneze - adaptace, které vznikají v embryích nebo larvách a přizpůsobují je vlastnostem jejich prostředí. U dospělých organismů není cenogeneze zachována. Příkladem koenogeneze jsou rohovinové útvary v tlamě larev bezocasých obojživelníků, které jim usnadňují potravu rostlinnou potravou. Během procesu metamorfózy v žábě mizí a trávicí systém je přebudován tak, aby se živil hmyzem a červy. Koenogeneze u amniot zahrnuje embryonální membrány, žloutkový váček a alantois a u placentárních savců a lidí také placentu a pupeční šňůru.
Cenogeneze, objevující se pouze v raných fázích ontogeneze, nemění typ organizace dospělého organismu, ale poskytuje vyšší pravděpodobnost přežití potomstva. Mohou být doprovázeny poklesem plodnosti a prodloužením embryonálního nebo larválního období, díky čemuž se organismus v postembryonálním nebo postlarválním období vývoje ukazuje jako zralejší a aktivnější. Poté, co vznikly a ukázaly se být užitečné, budou cenogeneze reprodukovány v následujících generacích. Amnion, který se poprvé objevil u předků plazů v období karbonu paleozoické éry, se tedy reprodukuje u všech obratlovců vyvíjejících se na souši, jak u vejcorodých plazů a ptáků, tak u placentárních savců.
Dalším typem fylogeneticky významných přeměn fylogeneze je fylembryogeneze. Představují odchylky od ontogeneze charakteristické pro předky, projevující se v embryogenezi, ale mající adaptační význam u dospělých forem. Vlasové pupeny se tedy objevují u savců ve velmi raných fázích embryonálního vývoje, ale samotné vlasy jsou důležité pouze u dospělých organismů.
Takové změny v ontogenezi, které jsou užitečné, jsou fixovány přirozeným výběrem a reprodukovány v následujících generacích. Tyto změny jsou založeny na stejných mechanismech, které způsobují vrozené vývojové vady: narušená proliferace buněk, pohyb, adheze, smrt nebo diferenciace. Od neřestí se však stejně jako cenogeneze odlišují svou adaptivní hodnotou, tzn. užitečnost a fixace přirozeným výběrem ve fylogenezi.
V závislosti na fázích embryogeneze a morfogeneze specifických struktur dochází k vývojovým změnám, které mají význam fylembryogeneze, rozlišují se tři typy.
1. anabolismus, nebo rozšíření, vznikají poté, co orgán prakticky dokončil svůj vývoj, a jsou vyjádřeny přidáním dalších fází, které mění konečný výsledek.
Mezi anabolismy patří takové jevy, jako je získání specifického tvaru těla u platýse až poté, co se z vajíčka vylíhne plůdek, k nerozeznání od jiných ryb, dále vzhled zakřivení páteře, splynutí švů v mozkové lebce a konečná redistribuce krevních cév v těle savců a lidí.
2. Odchylky - odchylky, které vznikají v procesu morfogeneze orgánů. Příkladem je vývoj srdce v ontogenezi savců, kdy rekapituluje trubicové stadium, dvoukomorovou a tříkomorovou stavbu, ale stadium tvorby neúplné přepážky, charakteristické pro plazy, je nahrazeno vývojem. přepážky, konstruované a umístěné odlišně a charakteristické pouze pro savce. Ve vývoji plic u savců se nachází i rekapitulace raných stádií předků a pozdější morfogeneze probíhá novým způsobem.
3. Archalaxis - změny, které jsou detekovány na úrovni primordií a jsou vyjádřeny porušením jejich rozdělení, časnou diferenciací nebo výskytem zásadně nových primordií. Klasickým příkladem archalaxis je
vývoj srsti u savců, k jejichž analizaci dochází ve velmi raných stádiích vývoje a od samého počátku se liší od anlage jiných kožních přívěsků obratlovců.
Podle typu archalaxis se u primitivních bezlebek objevuje notochord, u chrupavčitých ryb chrupavčitá páteř a u plazů se vyvíjejí nefrony sekundární ledviny.
Je zřejmé, že během evoluce vlivem anabolismu v ontogenezích potomků se plně realizuje základní biogenetický zákon, tzn. dochází k rekapitulacím všech vývojových stádií předků. V případě odchylek se rekapitulují raná stadia předků a pozdější jsou nahrazena vývojem novým směrem. Archalaxis zcela brání rekapitulaci ve vývoji těchto struktur a mění jejich samotné základy.
Porovnáme-li diagram fylembryogeneze s tabulkou K. Baera, která ilustruje zákon zárodečné podobnosti, je jasné, že Baer byl již velmi blízko objevu fylembryogeneze, ale absence evoluční myšlenky v jeho úvahách ano. nedovolit mu být o více než 100 let před vědeckým myšlením.
V evoluci ontogeneze se anabolismus nejčastěji vyskytuje jako fylembryogeneze, která jen v malé míře mění celý vývojový proces. Odchylky jako porušení morfogenetického procesu v embryogenezi jsou přirozeným výběrem často odmítány, a jsou proto mnohem méně časté. Archallaxe se v evoluci vyskytují jen zřídka, protože mění celý průběh embryogeneze, a pokud takové změny ovlivňují základy životně důležitých orgánů nebo orgánů, které jsou důležitými embryonálními organizačními centry, pak se často ukáží jako neslučitelné se životem.
Ve stejné fylogenetické skupině může v důsledku odlišné fylembryogeneze dojít k evoluci v různých orgánových systémech.
V ontogenezi savců jsou tedy sledována všechna stádia vývoje osového skeletu u subtypu obratlovců (anabolismus), ve vývoji srdce rekapitulují pouze raná stádia (odchylka), ve vývoji kožních přívěsků dochází vůbec žádná rekapitulace (archalaxis). Znalost typů fylembryogeneze v evoluci orgánových systémů strunatců je pro lékaře nezbytná k predikci možnosti výskytu vrozených vývojových vad atavistického charakteru u plodů a novorozenců. Pokud jsou totiž v orgánovém systému vyvíjejícím se anabolismem a deviací možné atavistické vývojové vady v důsledku rekapitulace rodových stavů, pak je to v případě archalaxy zcela vyloučeno.
Kromě cenogeneze a fylembryogeneze lze odchylky v době vzniku orgánu nalézt také ve vývoji ontogeneze - heterochronie - a místa jejich vývoje - heterotopie. První i druhý vedou ke změnám ve vzájemné korespondenci vyvíjejících se struktur a podléhají přísné kontrole přírodního výběru. Zachovány jsou pouze ty heterochronie a heterotopie, které se ukáží jako užitečné. Příkladem takových adaptivních heterochronií jsou posuny v době vzniku nejvitálnějších orgánů ve skupinách vyvíjejících se podle typu arogeneze. U savců a zejména u člověka tedy diferenciace předního mozku výrazně posouvá vývoj jeho ostatních částí.
Heterotopie vedou ke vzniku nových prostorových a funkčních spojení mezi orgány a zajišťují jejich další společný vývoj. Srdce, které se nachází pod hltanem ryb, tedy zajišťuje účinný průtok krve do žaberních tepen pro výměnu plynů. Přesun do retrosternální oblasti u suchozemských obratlovců se vyvíjí a funguje v jediném komplexu s novými dýchacími orgány - plícemi, a i zde plní především funkci dodávání krve do dýchacího systému k výměně plynů.
Heterochronie a heterotopie, v závislosti na tom, v jakých fázích embryogeneze a orgánové morfogeneze se objevují, mohou být považovány za fylembryogenezi různých typů. Pohyb rudimentů mozku vedoucí k jeho ohnutí, charakteristické pro amnioty a projevující se v počátečních fázích jeho diferenciace, je tedy archalaxie a heterotopie varlete u lidí z břišní dutiny přes tříselný kanál do šourek, pozorovaný na konci embryogeneze po jeho konečném vytvoření, - typická anabolie.
Někdy procesy heterotopie, identické ve výsledcích, mohou být fylembryogenezí různých typů. Například u různých tříd obratlovců je pohyb pletenců končetin velmi častý. U mnoha skupin ryb, které vedou životní styl u dna, jsou břišní ploutve (zadní končetiny) umístěny před prsními ploutvemi a u savců a lidí jsou ramenní pletence a přední končetiny v definitivním stavu umístěny výrazně kaudálně k místu ploutve. jejich prvotní formace. V tomto ohledu je inervace ramenního pletence v nich prováděna nervy spojenými nikoli s hrudními, ale s krčními segmenty míchy. U výše zmíněných ryb nejsou pánevní ploutve inervovány zadními kmenovými nervy, ale předními segmenty umístěnými před centry inervace prsních ploutví. To ukazuje na heterotopii při tvorbě ploutví již ve stádiu nejranějších rudimentů, zatímco k pohybu předního pletence končetin u člověka dochází v pozdějších stádiích, kdy je již plně dokončena jejich inervace. Je zřejmé, že v prvním případě heterotopie představuje archalaxii, zatímco ve druhém anabolii.
Cenogeneze, fylembryogeneze, stejně jako heterotopie a heterochronie, které se ukázaly být užitečné, jsou fixovány v potomstvu a reprodukovány v následujících generacích, dokud je nevytlačí nové adaptivní změny v ontogenezi a nenahradí je. Díky tomu ontogeneze nejen stručně opakuje evoluční cestu, kterou prošli naši předkové, ale také dláždí nové směry fylogeneze do budoucna.
Cenogeneze(z řeckého kainós - nový a ... genesis (Viz ... genesis) adaptace organismu, která se vyskytuje v embryonálním (fetálním) nebo larválním stádiu a nepřetrvává v dospělosti. Příkladem c. je savčí placenta, která zajišťuje plodu dýchání, výživu a vylučování; vnější žábry larev obojživelníků; vaječný zub u ptáků, který používají kuřata k proražení skořápky vajec; připojovací orgány u larev ascidiánů, plovoucí ocas u larev motolice - cerkárie atd. Termín „C.“ zavedl v roce 1866 E. Haeckel k označení těch vlastností, které narušují projevy palingeneze , tj. opakování vzdálených fází fylogeneze v procesu embryonálního vývoje jedince, neumožňují vysledovat sled fází fylogeneze jejich předků během ontogeneze moderních forem, tedy porušují biogenetický zákon. Na konci 19. stol. Každá změna v průběhu ontogeneze charakteristická pro předky se začala nazývat barva (němečtí vědci E. Mehnert, F. Keibel a další). Moderní chápání pojmu „C“. vznikla jako výsledek práce A. N. Severtsova, který si pro tento koncept ponechal pouze význam provizorních zařízení, neboli embryo-adaptací. Viz také Filembryogeneze.
Cenogeneze(řecký kainos new + genesis původ, vznik) - výskyt adaptací v embryu nebo larvě na životní podmínky, které nejsou charakteristické například pro dospělá stádia. tvorba membrán v embryích vyšších živočichů.
nebo tak
FYLEMBRYOGENEZE (z řeckého phylon - rod, kmen, embryo - embryo a genesis - původ), evoluční změna v ontogenezi orgánů, tkání a buněk, spojená jak s progresivním vývojem, tak s redukcí. Doktrínu fylembryogeneze vypracoval ruský evoluční biolog A.N. Severtsov. Způsoby (metody) fylembryogeneze se liší dobou výskytu během vývoje těchto struktur.
Pokračuje-li vývoj určitého orgánu u potomků po stádiu, ve kterém skončil u předků, nastává anabolie (z řeckého anabole – vzestup) – prodloužení konečného stádia vývoje. Příkladem je vznik čtyřkomorového srdce u savců. Obojživelníci mají tříkomorové srdce: dvě síně a jednu komoru. U plazů vzniká v komoře přepážka (první anabolie), u většiny z nich je však tato přepážka neúplná – pouze omezuje promíchávání arteriální a venózní krve. U krokodýlů a savců pokračuje vývoj septa až do úplného oddělení pravé a levé komory (druhá anabolie). U dětí je někdy jako atavismus nedostatečně vyvinutá interventrikulární přepážka, což vede k vážnému onemocnění vyžadujícímu chirurgický zákrok.
Prodloužení vývoje orgánu nevyžaduje hluboké změny v předchozích fázích jeho ontogeneze, proto je anabolismus nejčastější metodou fylembryogeneze. Fáze vývoje orgánu předcházející anabolismu zůstávají srovnatelné se stádii rodové fylogeneze (tj. jsou to rekapitulace) a mohou sloužit k jeho rekonstrukci (viz Biogenetický zákon). Pokud se vývoj orgánu v mezistupních odchýlí od cesty, po které probíhala jeho ontogeneze u jeho předků, nastává deviace (z pozdně latinského deviatio - odchylka). Například u ryb a plazů se šupiny objevují jako ztluštění epidermis a podkladové vrstvy pojivové tkáně kůže – koria. Postupným zahušťováním se tato analage ohýbá směrem ven. Poté u ryb korium zkostnatí, tvořící se kostní šupiny prorazí epidermis a přesunou se na povrch těla. Naopak u plazů se kost netvoří, ale epidermis keratinizuje a tvoří zrohovatělé šupiny ještěrek a hadů. U krokodýlů může korium osifikovat a tvoří kostěný základ rohovitých šupin. Odchylky vedou k hlubší restrukturalizaci ontogeneze než anabolismus, takže jsou méně časté.
Nejméně často dochází ke změnám primárních rudimentů orgánů – archalaxis (z řeckého arche – začátek a allaxis – změna). V případě odchylky lze rekapitulaci vysledovat od vzniku orgánu až po okamžik vývojové odchylky. V archalaxii nedochází k rekapitulaci. Příkladem je vývoj obratlových těl u obojživelníků. U fosilních obojživelníků - stegocefalií a u moderních bezocasých obojživelníků jsou obratlová těla vytvořena kolem tětivy několika, obvykle tří na každé straně těla, samostatných anlage, které se pak spojí a vytvoří tělo obratle. U ocasatých obojživelníků se tato anlage neobjevuje. Osifikace roste nahoře a dole a pokrývá notochord, takže se okamžitě vytvoří kostní trubice, která se ztluštěním stává tělem obratle. Tato archallaxa je důvodem stále diskutované otázky původu ocasatých obojživelníků. Někteří vědci se domnívají, že pocházejí přímo z lalokoploutvých ryb, bez ohledu na ostatní suchozemské obratlovce. Jiní říkají, že ocasatí obojživelníci se velmi brzy odchýlili od ostatních obojživelníků. Ještě další, opomíjející vývoj obratlů, dokazují blízkou příbuznost ocasatých a bezocasých obojživelníků.
K redukci orgánů, které ztratily svůj adaptační význam, dochází i fylembryogenezí, především negativním anabolismem – ztrátou konečných vývojových fází. V tomto případě se orgán buď nevyvine a stane se rudimentem, nebo projde zpětným vývojem a zcela zmizí. Příkladem rudimentu je lidské slepé střevo - nedostatečně vyvinuté slepé střevo, příkladem úplného zmizení je ocas žabích pulců. Po celou dobu života ve vodě ocas roste, na jeho konci přibývají nové obratle a svalové segmenty. Během metamorfózy, kdy se pulec promění v žábu, se ocas rozpustí a proces probíhá v opačném pořadí - od konce k základně. Fylembryogeneze je hlavní metodou adaptačních změn ve struktuře organismů během fylogeneze.
Lístek 97
Principy (metody) fylogenetických přeměn orgánů a funkcí. Soulad mezi strukturou a funkcí v živých systémech. Multifunkčnost Kvantitativní a kvalitativní změny funkcí biologických struktur.
.
Principy fylogenetických přeměn
Autorita nazýváme historicky vyvinutý specializovaný systém tkání, vyznačující se vymezením, stálostí tvaru, lokalizací, vnitřní stavbou oběhových a inervačních drah, vývojem v ontogenezi a specifickými funkcemi. Struktura orgánů je často velmi složitá. Většina z nich je multifunkčních, tzn. plní několik funkcí současně. Současně se různé orgány mohou podílet na provádění jakékoli komplexní funkce Skupina orgánů podobného původu, které se spojují, aby vykonávaly komplexní funkci, se nazývá Systém(oběhový, vylučovací apod.) Pokud stejnou funkci plní skupina orgánů různého původu, jedná se o tzv. zařízení. Příkladem je dýchací aparát, který se skládá jak ze samotných dýchacích orgánů, tak z prvků kosterního a svalového systému, které zajišťují dýchací pohyby.
V procesu ontogeneze dochází k vývoji a často i náhradě některých orgánů jinými. Orgány zralého organismu se nazývají definitivní; orgány, které se vyvíjejí a fungují pouze v embryonálním nebo larválním vývoji - prozatímní. Příklady provizorních orgánů jsou žábry larev obojživelníků, primární ledvina a embryonální membrány vyšších obratlovců (amniota).
V historickém vývoji mohou mít orgánové přeměny progresivní nebo regresivní charakter. V prvním případě se orgány zvětšují a stávají se složitější strukturou, ve druhém se zmenšují a jejich struktura se zjednodušuje.
Pokud dva organismy umístěné na různých úrovních organizace mají orgány, které jsou postaveny podle jediného plánu, umístěné na stejném místě a vyvíjejí se podobným způsobem ze stejných embryonálních základů, pak to naznačuje vztah těchto organismů. Takové orgány se nazývají homologní. Homologní orgány často plní stejnou funkci (například srdce ryb, obojživelníků, plazů a savců), ale v procesu evoluce se funkce mohou měnit (například přední končetiny ryb a obojživelníků, plazů a ptáků).
Když nepříbuzné organismy žijí ve stejném prostředí, mohou se u nich vyvinout podobné adaptace, které se projeví ve vzhledu podobný orgány. Podobné orgány plní stejné funkce, ale jejich struktura, umístění a vývoj jsou ostře odlišné. Příklady takových orgánů jsou křídla hmyzu a ptáků, končetiny a čelistní aparát členovců a obratlovců.
Struktura orgánů přesně odpovídá funkcím, které vykonávají. Navíc v historických proměnách orgánů je změna funkcí jistě doprovázena změnou morfologické charakteristiky orgánu.
Základním principem evoluce organických struktur je princip diferenciace. Diferenciace je rozdělení homogenní struktury na samostatné části, které v důsledku různých poloh, spojení s jinými orgány a různých funkcí získávají specifickou strukturu. Komplikace konstrukce je tedy vždy spojena s komplikací funkcí a specializací jednotlivých částí. Diferencovaná struktura plní několik funkcí a její struktura je složitá.
Příkladem fylogenetické diferenciace může být evoluce oběhového systému v kmeni strunatců. U zástupců bezlebkového podtypu je tedy strukturován velmi jednoduše: jeden kruh krevního oběhu, absence srdce a kapilár v systému žaberních tepen.
Nadtřída ryb má dvoukomorové srdce a žaberní kapiláry. U obojživelníků je oběhový systém poprvé rozdělen do dvou oběhových kruhů a srdce se stává tříkomorovým. Maximální diferenciace je charakteristická pro oběhový systém savců, jejichž srdce je čtyřkomorové a v cévách je dosaženo úplného oddělení žilního a arteriálního krevního toku.
Jednotlivé části diferencující, dříve homogenní struktury, specializované na plnění jedné funkce, se stávají funkčně stále více závislé na jiných částech této struktury a na organismu jako celku. Tato funkční podřízenost jednotlivých složek systému v celém organismu se nazývá integrace.
Čtyřkomorové srdce savců je příkladem vysoce integrované struktury: každé oddělení plní pouze svou speciální funkci, která nemá žádný význam izolovaně od funkcí ostatních oddělení. Proto je srdce vybaveno autonomním systémem funkční regulace v podobě parasympatického atrioventrikulárního nervového uzlu a zároveň je přísně podřízeno neurohumorálnímu regulačnímu systému těla jako celku.
Současně s diferenciací se tedy sleduje i podřízení částí ucelenému systému těla, tzn. integrační proces.
V procesu evoluce je přirozené, že vznik nové struktury a jejich zmizení. Je založena na principu diferenciace, která se projevuje na pozadí primární polyfunkčnosti a schopnosti funkcí se kvantitativně měnit. Jakákoli struktura v tomto případě vzniká na základě předchozích struktur, bez ohledu na to, na jaké úrovni organizace živých věcí proces fylogeneze probíhá. Je tedy známo, že přibližně před 1 miliardou let se původní protein globin po zdvojení původního genu diferencoval na myo- a hemoglobin - proteiny, které jsou součástí svalových a krevních buněk, a proto se diferencují podle funkce. Ve fylogenezi centrálního nervového systému strunatců lze také vidět diferenciaci a změnu funkcí struktur: mozek se tvoří z předního konce neurální trubice. Stejně tak vznikají nové biologické druhy ve formě izolovaných populací původních druhů (viz § 11.6) a nové biogeocenózy - v důsledku diferenciace již existujících (viz § 16.2).
Vzhledem k tomu, že fylogeneze konkrétních orgánových systémů budou zvažovány níže, zastavíme se podrobněji u vzorců vzhledu a mizení orgánů. Příklad vznik orgánů je původem dělohy placentárních savců z párových vejcovodů. S prodlužováním embryonálního vývoje savců vyvstává potřeba delšího udržení embrya v těle matky. To lze provést pouze v kaudálních úsecích vejcovodů, jejichž dutina je zvětšena a stěna je diferencována tak, že je na ni připojena placenta, čímž je zajištěn vztah mezi matkou a plodem. V procesu přirozeného výběru se zachovali a úspěšně rozmnožili především ti savci, v jejichž tělech se potomci vyvíjeli nejdéle. V důsledku toho vznikl nový orgán - děloha, která poskytuje embryu optimální podmínky pro nitroděložní vývoj (viz kapitola 14.5.3) a zvyšuje míru přežití odpovídajícího druhu.
Při vzniku tak složitějšího a specializovanějšího orgánu, jako je oko, jsou pozorovány stejné vzorce. Tvorba zrakového orgánu, stejně jako všech smyslových orgánů, je založena na kožních epiteliálních buňkách, mezi nimiž se rozlišují receptorové buňky, zejména fotosenzitivní buňky. Jejich spojení do skupin vede ke vzniku primitivních oddělených orgánů zraku, což umožňuje zvířatům pouze hodnotit osvětlení. Ponoření takového orgánu citlivého na světlo pod kůži zajišťuje bezpečnost jemných buněk, ale vizuální funkce může být dosažena pouze díky vzhledu průhlednosti integumentu. Citlivost primitivního zrakového orgánu na světlo se zvyšuje se ztluštěním průhledných krytů a jejich získáním schopnosti lámat světlo a soustředit jeho paprsky na citlivé buňky oka. Složitý orgán vyžaduje pomocný aparát - ochranné struktury, svaly, které jej pohybují atd. Zvýšená úroveň složitosti organizace oka je nutně doprovázena komplikací regulace jeho funkcí, což se projevuje posílením jeho integrace jako integrálního systému.
Zmizení nebo redukce orgánu ve fylogenezi může být spojena se třemi různými důvody a má různé mechanismy. Za prvé, orgán, který dříve vykonával důležité funkce, se může v nových podmínkách ukázat jako škodlivý. Proti tomu působí přírodní výběr a orgán může zcela rychle zmizet. Existuje jen málo příkladů takového přímého zmizení orgánů. Mnoho hmyzu malých oceánských ostrovů je tedy bezkřídlých kvůli neustálému vyřazování létajících jedinců z jejich populací větrem. Zánik orgánů je častěji pozorován v důsledku jejich náhrady novými strukturami, které plní stejné funkce s větší intenzitou. Tak např. u plazů a savců mizí přední pupeny a primární ledviny, které jsou funkčně nahrazeny ledvinami sekundárními. Stejně tak u ryb a obojživelníků je notochord přemístěn páteří.
Nejčastější cesta k zániku orgánů vede přes postupné oslabování jejich funkcí. Takové situace obvykle nastávají při změně životních podmínek. Orgán, který neplní téměř žádné funkce, se vymyká kontrole přirozeného výběru a obvykle vykazuje zvýšenou variabilitu. Výsledné změny způsobují narušení korelačních spojení s ostatními částmi těla. Kvůli tomu se takový orgán často stává škodlivým a přírodní výběr proti němu začíná působit.
V lékařské praxi je všeobecně známo, že rudimentární orgány u lidí se vyznačují širokou variabilitou. Například třetí velké stoličky neboli „zuby moudrosti“ se vyznačují nejen výraznou variabilitou ve struktuře a velikosti, ale také různými obdobími erupce a také zvláštní náchylností ke vzniku zubního kazu. Někdy nevybuchnou vůbec a často, když vybuchnou, jsou během několika příštích let zcela zničeny. Totéž platí pro červovité slepé střevo (apendix), které může mít za normálních okolností délku od 2 do 20 cm a může být umístěno různě (za pobřišnicí, na dlouhém mezenteriu, za slepým střevem atd.). Zánět slepého střeva (apendicitida) se navíc vyskytuje mnohem častěji než zánětlivé procesy v jiných částech střeva.
Proces redukce orgánu je opačný k jeho normální morfogenezi. Nejprve vypadnou záložky takových částí varhan, které se běžně tvoří jako poslední. Když jsou končetiny člověka nedostatečně vyvinuté, falangy 1. a 5. prstu, které se vyvíjejí jako poslední, jsou obvykle jako první nedostatečně vyvinuté. U kytovců, kteří jsou zcela bez zadních končetin kvůli oslabení jejich funkcí ve fylogenezi, stále zůstávají rudimenty prvků pánevního pletence, které se tvoří velmi brzy v procesu morfogeneze.
Studie genetického základu orgánové redukce prokázaly, že strukturální geny regulující morfogenezi nezanikají, zatímco geny regulující dobu vzniku rudimentárních orgánů, případně geny odpovědné za fenomén induktivních interakcí ve vyvíjejícím se embryu, procházejí významnými změnami. Když je totiž mezodermální materiál dna ústní dutiny ještěrčího embrya transplantován do ústní dutiny vyvíjejícího se kuřátka, je možné, aby toto kuřátko vytvořilo zuby typické struktury a transplantace kožního mezodermu ještěrka pod epidermis hřbetu kuřátka vede k tvorbě typických zrohovatělých šupin v něm namísto peří.
Ontogeneze je individuální vývoj, komplex vývojových procesů jedince, zejména od vzniku zygoty až po smrt. K vývoji dochází díky implementaci genetické informace získané od rodičů. Na jeho realizaci mají významný vliv podmínky prostředí. Fylogeneze je historický vývoj druhu, evoluční vývoj organismů. Oba procesy spolu úzce souvisí. Se znalostí směrů a proměn orgánů a jejich systémů v procesu historického vývoje je možné porozumět a vysvětlit vývojové anomálie, které vznikají během embryogeneze.
Souvislost mezi otnogenezí a fylogenezí se odráží v řadě biologických zákonů a zákonitostí.V roce 1828 formuloval Karl Baer tři zákony:
1. Zákon zárodečné podobnosti - embryo jakéhokoli vyššího zvířete není podobné jinému zvířeti, ale podobné jeho zárodku
2. zákon sekvenčního vzhledu znaků - obecnější znaky charakteristické pro danou velkou skupinu zvířat se v jejich embryích odhalují dříve než specializovanější znaky
3. zákon mebryonální divergence - každé embryo dané formy živočichů neprochází jinými formami, ale postupně se od nich odděluje.
Tyto zákony lze vysvětlit tak, že v raných fázích embryogeneze jsou si embrya zvířat různých tříd obratlovců (například ryb, ptáků, savců) navzájem podobná. Postupem času se mezi nimi objevují rozdíly v rámci tříd a poté v rámci řádů (příklad: prasečí a lidská embrya)
K. Baerův zákon zárodečné podobnosti
V roce 1828 Karl von Baer formuloval vzorec nazvaný Baerův zákon: „Čím dřívější fáze individuálního vývoje jsou porovnávány, tím více podobností lze nalézt. Při porovnání fází vývoje embryí různých druhů a tříd strunatců učinil K. Baer následující závěry.
Embrya zvířat stejného typu jsou v raných fázích vývoje podobná.
Důsledně se pohybují ve svém vývoji od obecnějších charakteristik typu ke stále specifičtějším. Nakonec se vyvinou znaky, které naznačují, že embryo patří k určitému rodu, druhu a nakonec k jednotlivým znakům.
Embrya různých zástupců stejného typu se postupně od sebe oddělují.
Následný vývoj evoluční představy umožnil vysvětlit podobnost raných embryí jejich historickou příbuzností a jejich nabýváním stále specifičtějších rysů s postupnou vzájemnou izolací - skutečnou izolací odpovídajících tříd, řádů, čeledí, rody a druhy v procesu evoluce.
Embryonální podobnost je nyní vysvětlována skutečným vztahem organismů a jejich postupná divergence (embryonální divergence) slouží jako zřejmý odraz historické divergence těchto forem (fylogenetická divergence). V důsledku toho lze historii daného druhu sledovat individuálním vývojem.
Cenogeneze - změny v ontogenezi vedoucí k vybočení z cesty rodových forem, adaptace, které vznikají u embryí a larev, adaptace na jejich prostředí. U dospělých organismů není cenogeneze zachována, tzn. projevující se až v raných fázích ontogeneze, nemění typ organizace dospělého organismu, ale poskytují vyšší stupeň přežití potomstva. Například u hobitů jako plodových organismů zahrnuje cenogeneze zárodečné blány, žloutkový váček a alantois, u hobitů jako placentárních živočichů také placentu s pupeční šňůrou.
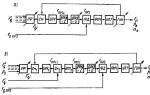
Fylembryogeneze jsou embryonální novotvary, které mají fylogenetický význam. Doba jejich vzniku a metody jsou různé (vyvíjí se příklad ze žraločích šupin: 1- zrohovatělá štípance plaza odchylkou 2 ptačí peří - anabolie 3 - chlupy savce - archalaxis. Evoluce je častěji anabolií, proto je pozorována rekapitulace Doba vzniku fylembryogeneze: 1 počáteční stadia morfogeneze (archalaxe - vývoj orgánu se zcela mění. Nedochází k rekapitulaci. 2 - střední stadia (odchylka - odchylka ve vývoji) 3 - konečná stadia (anabolie - přetlak ve vývoji orgánu)
Člověk a biosféra. Noosféra je nejvyšším stupněm vývoje biosféry. Reakce těla na změny v okolní situaci. Příklady. Liebig-Tienemannovo pravidlo. Liebigův sud. Princip Le Chatelier-Brown.
Biosféra a člověk. Moderní člověk vznikl asi před 30-40 tisíci lety. Od té doby začal ve vývoji biosféry působit nový antropogenní faktor. První kultura vytvořená člověkem, paleolit (doba kamenná), trvala přibližně 20-30 tisíc let; se shodovalo s dlouhým obdobím zalednění. Ekonomickým základem lidské společnosti byl lov velkých zvířat: červených a sobů, nosorožců srstnatých, osla, koně, mamuta, zubra. Na lidských místech doby kamenné se nalézají četné kosti divokých zvířat jako důkaz úspěšného lovu. Intenzivní hubení velkých býložravců vedlo k poměrně rychlému snížení jejich počtu a vyhynutí mnoha druhů. Pokud mohli malí býložravci díky své vysoké porodnosti nahradit ztráty z pronásledování lovci, velká zvířata byla díky evoluční historii o tuto příležitost připravena. Další potíže pro býložravce vznikly v důsledku změn přírodních podmínek na konci paleolitu. Před 10-13 tisíci lety došlo k prudkému oteplení, ledovec ustoupil, po Evropě se rozšířily lesy a vyhynula velká zvířata. To vytvořilo nové životní podmínky a zničilo dosavadní ekonomickou základnu lidské společnosti. Skončilo období jeho vývoje, charakterizované pouze používáním potravin, tzn. čistě spotřebitelský postoj k životnímu prostředí. V další éře neolitu, spolu s lovem (kůň, divoké ovce, jelen, divoká prasata, bizon atd.), rybolovem a sběrem (měkkýš, ořechy, bobule, ovoce) nabýval na významu proces výroby potravin. Probíhají první pokusy o domestikaci zvířat a šlechtění rostlin a začíná výroba keramiky. Již před 9-10 tisíci lety existovaly osady, mezi jejichž zbytky byla nalezena pšenice, ječmen, čočka, kosti domácích zvířat, koz, ovcí a prasat. Na různých místech západní a střední Asie, na Kavkaze a v jižní Evropě se rozvíjejí základy zemědělství a chovu dobytka. Oheň je široce používán jak k ničení vegetace v zemědělství sekát a spálit, tak jako prostředek k lovu. Začíná rozvoj nerostných surovin a rodí se hutnictví. Populační růst a kvalitativní skok ve vývoji vědy a techniky za poslední dvě století, zejména dnes, vedly k tomu, že lidská činnost se stala faktorem v planetárním měřítku, vůdčí silou dalšího vývoje biosféry. V A. Vernadsky věřil, že vliv vědeckého myšlení a lidské práce určil přechod biosféry do nového stavu - noosféry (sféry mysli).
NOOSFÉRA – NEJVYŠŠÍ ETAPA VÝVOJE BIOSFÉRY
Sféra interakce mezi společností a přírodou, v rámci které rozumné
činnost se jeví jako hlavní určující faktor rozvoje biosféry a
lidstvo se nazývá noosféra.
Termín „noosféra“ byl poprvé použit v letech 1926–1927. používané francouzskými vědci E.
Lecroix (1870 – 1954) a P. Teilhard de Chardin (1881 – 1955) ve smyslu „nové
obal“, „vrstva myšlení“, která vznikla na konci třetihor,
se odvíjí mimo biosféru nad světem rostlin a zvířat. V jejich
noosféra je ideální, duchovní ("myšlení") skořápka Země,
vznikly s příchodem a rozvojem lidského vědomí. Přednost plnění
Tento koncept svým materialistickým obsahem patří akademikovi V.I.
Vernadsky (1965, 1978).
Podle V.I. Vernadského je člověk součástí živé hmoty,
podléhá obecnému zákonu o organizaci biosféry, mimo ni
nemůže existovat. Člověk je součástí biosféry, tvrdil vynikající
vědec. Cílem sociálního rozvoje by mělo být udržení organizace
biosféra. Zachování jeho primární organizace – „intaktní
příroda“ – nenese v sobě tvůrčí princip do mocné geologické síly.“ „A
před ním, před jeho myšlením a prací, vyvstává otázka restrukturalizace biosféry
v zájmu svobodně smýšlejícího lidstva jako celku. To je nové
stav biosféry, ke kterému se blížíme, aniž bychom si toho všimli, je
„noosféra“. Noosféra představuje kvalitativně novou etapu evoluce
biosféra, ve které se jako nové vytvářejí nové formy její organizace
jednota vyplývající ze vzájemného působení přírody a společnosti. V něm
přírodní zákony jsou úzce propojeny se socioekonomickými zákony
rozvoj společnosti, tvořící nejvyšší hmotnou celistvost „polidštěných
Příroda."
V. I. Vernadského, který předpověděl nástup éry vědecké a technické
revoluce ve 20. století, hlavní předpoklad přechodu biosféry na noosféru
považován za vědecké myšlení. Jeho hmotné vyjádření v člověkem přeměněném
biosféra je práce. Jednota myšlení a práce nevytváří pouze nové
sociální podstatu člověka, ale také předurčuje přechod biosféry do
noosféra. „Věda je maximální síla k vytvoření noosféry“ - to je hlavní věc
postavení V. I. Vernadského v doktríně biosféry, která volá po transformaci,
a nezničit ekuménu.
Reakce těla na změny v okolní situaci.
Stejný faktor může mít optimální účinek na různé organismy při různých hodnotách. Kromě toho se živé organismy dělí na ty, které jsou schopny existovat v širokém nebo úzkém rozsahu změn jakéhokoli faktoru prostředí. Organismy se přizpůsobují každému faktoru prostředí relativně nezávisle. Organismus může být adaptován na úzký rozsah jednoho faktoru a široký rozsah jiného. Pro tělo je důležitá nejen amplituda, ale i rychlost kmitání konkrétního faktoru.
Pokud vliv podmínek prostředí nedosahuje extrémních hodnot, živé organismy na něj reagují určitými akcemi nebo změnami svého stavu, což v konečném důsledku vede k přežití druhu. Překonání nepříznivých účinků zvířat je možné dvěma způsoby:
1) tím, že se jim vyhýbáte,
2) získáním vytrvalosti. Základem reakcí rostlin je rozvoj adaptačních změn v jejich struktuře a životních procesech. Variabilita je jednou z hlavních vlastností živých věcí na různých úrovních jejich organizace. Genetická variabilita je základem dědičné variability znaků.
Liebigovo minimální pravidlo
Podle Liebigova pravidla minima je sklizeň (produkce), její velikost a stabilita v čase řízena zdrojovou proměnnou, jako je prostor, čas, hmota, energie a diverzita, kterých je minimum. V současné době je toto pravidlo rozšířeno na fungování různých objektů a obecně říká, že stav funkce je určen faktorem, který má minimální hodnotu. V interpretaci Yu Odum (1986) na úrovni organismu ve stacionárním stavu bude limitující látkou ta vitální látka, jejíž dostupné množství se nejvíce blíží požadovanému minimu. E. A. Mitscherlich formuloval pravidlo kombinovaného působení faktorů, které objasňuje Liebigovo minimální pravidlo. Pro zvířata jsou limitujícími faktory zpravidla: dostupnost dostatečného množství potravy, vhodné úkryty (úkryty), klimatické podmínky.
Zákon zárodečné podobnosti. Výzkumy na počátku 19. století začaly poprvé upozorňovat na podobnost raných fází vývoje embryí vyšších živočichů se stádii organizační složitosti vedoucí od nízko organizovaných forem k progresivním. Porovnání fází vývoje embryí různých druhů a tříd strunatců, K. Baer učinil následující závěry:
1. Embrya zvířat stejného typu jsou v raných fázích vývoje podobná.
2. Důsledně se pohybují ve svém vývoji od obecnějších charakteristik typu ke stále specifičtějším. Nakonec se vyvinou znaky, které naznačují, že embryo patří k určitému rodu, druhu a nakonec k jednotlivým znakům.
3. Postupně se od sebe oddělují embrya různých zástupců stejného typu.
K. Baer, který nebyl evolucionistou, nedokázal spojit vzorce individuálního vývoje, které objevil, s procesem fylogeneze. Proto zobecnění, která učinil, měla význam pouze empirických pravidel.
Následný vývoj evoluční myšlenky umožnil vysvětlit podobnost raných embryí jejich historickou příbuzností a jejich osvojováním stále specifičtějších znaků s postupnou vzájemnou izolací - skutečnou izolací odpovídajících tříd, řádů, rodin, rody a druhy v procesu evoluce.
Brzy po objevu zákona embryonální podobnosti Charles Darwin ukázal, že tento zákon svědčí o společném původu a jednotě počátečních fází evoluce v rámci typu.
Ontogeneze je opakováním fylogeneze. Srovnáním ontogeneze korýšů s morfologií jejich vyhynulých předků dospěl F. Muller k závěru, že žijící korýši ve svém vývoji opakují cestu, kterou urazili jejich předci. Transformace ontogeneze v evoluci se podle Mullera provádí díky jejímu prodloužení tím, že se k ní přidávají další stupně nebo extenze. Na základě těchto pozorování, jakož i studia vývoje strunatců, formuloval E. Haeckel (1866) základní biogenetický zákon, podle kterého ontogeneze představuje stručnou a rychlou rekapitulaci fylogeneze.
Opakování struktur charakteristických pro předky v embryogenezi potomků se nazývá rekapitulace. Rekapitulujte nejen morfologické znaky - notochord, analáž žaberních štěrbin a žaberních oblouků u všech strunatců, ale i znaky biochemické organizace a fyziologie. Při evoluci obratlovců tedy dochází k postupné ztrátě enzymů nezbytných pro odbourávání kyseliny močové, produktu metabolismu purinů. U většiny bezobratlých je konečným produktem odbourávání kyseliny močové amoniak, u obojživelníků a ryb močovina, u mnoha plazů alantoin a u některých savců se kyselina močová vůbec nerozkládá a je vylučována močí. V embryogenezi savců a lidí jsou zaznamenány biologické a biochemické rekapitulace: uvolňování amoniaku ranými embryi, později močoviny, pak alantoinu a v posledních fázích vývoje - kyseliny močové.
V ontogenezi vysoce organizovaných organismů však není vždy dodržováno striktní opakování etap historického vývoje, jak vyplývá z biogenetického zákona. Lidské embryo tedy nikdy neopakuje dospělá stádia ryb, obojživelníků a plazů savců, ale je v řadě znaků podobné pouze jejich embryím. Raná stádia vývoje zůstávají nejkonzervativnější, díky čemuž rekapitulují úplněji než ty pozdější. To je způsobeno skutečností, že jedním z nejdůležitějších mechanismů pro integraci raných fází embryogeneze je embryonální indukce a struktury embrya, které se tvoří jako první, jako je notochord, nervová trubice, hltan, střevo a somity, jsou organizační centra embrya na kterých závisí.celý průběh vývoje.
Genetický základ rekapitulací spočívá v jednotě mechanismů genetického řízení vývoje, která je zachována na základě společných genů pro regulaci ontogeneze, které dědí příbuzné skupiny organismů po společných předcích.
Ontogeneze jako základ fylogeneze. Doktrína A. N. Severtsova o fylembryogenezi. Anabolie, deviace a archalaxie. Heterochronie a heterotopie biologických struktur ve vývoji ontogeneze.
Spoléháme-li se pouze na základní biogenetický zákon, je nemožné vysvětlit proces evoluce: nekonečné opakování toho, co bylo předáno, samo o sobě nezrodí něco nového. Život na Zemi tedy existuje díky střídání generací konkrétních organismů, jeho evoluce probíhá díky změnám v jejich ontogenezích. Tyto změny se scvrkávaly na skutečnost, že specifické ontogeneze se odchylují od cesty vytyčené rodovými formami a získávají nové rysy.
Mezi takové odchylky patří například koenogeneze – adaptace, které vznikají u embryí nebo larev a přizpůsobují je vlastnostem prostředí. U dospělých organismů není cenogeneze zachována. Příkladem koenogeneze jsou rohovinové útvary v tlamě larev bezocasých obojživelníků, které jim usnadňují potravu rostlinnou potravou. Během procesu metamorfózy v žábě mizí a trávicí systém je přestavěn tak, aby se živil hmyzem a červy. Koenogeneze u amniot zahrnuje embryonální membrány, žloutkový váček a alantois a u placentárních savců a lidí také placentu a pupeční šňůru.
Cenogeneze, objevující se pouze v raných fázích ontogeneze, nemění typ organizace dospělého organismu, ale poskytuje vyšší pravděpodobnost přežití potomstva. Mohou být doprovázeny poklesem plodnosti a prodloužením embryonálního nebo larválního období, díky čemuž se organismus v postembryonálním a postlarválním období vývoje ukazuje jako zralejší a aktivnější. Poté, co vznikla a ukázala se užitečná, bude cenogeneze reprodukována v následujících generacích. Amnion, který se poprvé objevil u předků plazů v období karbonu paleozoické éry, se tedy reprodukuje u všech obratlovců vyvíjejících se na souši, jak u vejcorodých plazů a ptáků, tak u placentárních savců.
Dalším typem fylogeneticky významných přeměn fylogeneze je fylembryogeneze. Představují odchylky od ontogeneze charakteristické pro předky, objevující se v embryogenezi, ale mající adaptační význam u dospělých forem. Vlasové pupeny se tedy objevují u savců ve velmi raných fázích embryonálního vývoje, ale samotné vlasy jsou důležité pouze u dospělých organismů. (fylembryogeneze je komplexem dědičně podmíněných adaptačních přeměn ontogeneze).
Takové změny v ontogenezi, které jsou užitečné, jsou fixovány přirozeným výběrem a reprodukovány v následujících generacích. Tyto změny jsou založeny na stejných mechanismech, které způsobují vrozené vývojové vady: poruchy proliferace buněk, jejich pohyb, adheze, diferenciace nebo smrt. Co je však odlišuje od neřestí, stejně jako od cenogeneze, je jejich adaptivní hodnota, ona užitečnost a fixace přirozeným výběrem ve fylogenezi.
V závislosti na fázích ontogeneze a morfogeneze konkrétních struktur dochází k vývojovým změnám, které mají význam fylembryogeneze, rozlišují se na tři typy.
1. Anabolismus nebo extenze nastávají poté, co orgán prakticky dokončil svůj vývoj, a jsou vyjádřeny přidáním dalších fází, které mění konečný výsledek.
Mezi anabolismy patří takové jevy, jako je získání specifického tvaru těla u platýse až poté, co se z vajíčka vylíhne plůdek, k nerozeznání od jiných ryb, dále vzhled zakřivení páteře, splynutí švů mozkové lebky, konečné přerozdělení krevních cév v těle savců a lidí, prsní ploutve u náručníku (Trigla) se nejprve vyvíjejí jako ostatní druhy ryb a poté dochází k anabolii - přední tři paprsky ploutve rostou jako prstovité přívěsky. Plodnice hub mají nástavec pro lepší distribuci výtrusů. Ledviny vyšších obratlovců (předledvinové, primární a sekundární).
V evoluci prostřednictvím anabolie se předchozí, dříve předkové konečné stádium vývoje ukazuje jako palingenetická rekapitulace. Odráží předchozí stupeň evoluce, organizaci předků. E. Haeckel nazval palingenezi charakteristikou embrya, rekapitulační charakteristikou předků.
2. Odchylky, resp. vyhýbání se vývoji mezistupňů ontogeneze (změna dráhy).
Příkladem odchylky je vývoj šupin u žraločích ryb a plazů. U žraloků, stejně jako u plazů, se nejprve objevují ztluštění epidermis, pod kterými se hromadí buňky pojivové tkáně koria. Tato analage rostou směrem k povrchu těla, postupně mají podobu šupin. U žraloků slouží buňky pojivové tkáně koria jako základ pro rozvoj osifikace a tvorbu kostních šupin, které perforují epidermis. U plazů se naopak tvoří šupiny v důsledku keratinizace epidermálních záhybů. Je pravděpodobné, že rostlinné hlízy a cibule vznikly odchylkou od primárního embryonálního pupenu. Vývoj srdce v ontogenezi savců, v níž rekapituluje trubicové stadium, dvoukomorovou, tříkomorovou stavbu, ale pro stadium tvorby neúplné přepážky, charakteristické pro plazy, nahrazuje vývoj přepážka, konstruovaná a umístěná odlišně a charakteristická pouze pro savce. Ve vývoji plic u savců se nachází i rekapitulace raných stádií předků, pozdější morfogeneze jde jinou cestou.
3. Archalaxis neboli změna primárních primordií. Zároveň se mění analage orgánu a vývoj od samého počátku jde na rozdíl od svých předků jinou cestou.
Například vývoj páteře u hadů ve srovnání s ještěry. Počet obratlů u ještěrek se pohybuje od 30 do 35, u velkých hadů dosahuje 500 a více. Prodlužování těla u hadů nastává v důsledku kladení dalších somitů, včetně segmentu svalů a obratlů. Ve stejném stádiu vývoje (soudím podle srovnání hlav) se u gekona (ještěrka) tvoří 24 somitů, u užovky užovky 34. V další fázi se u gekona vyvíjí 42 somitů a u hada 142. Další obratle užovky ve srovnání s obratli geckoho homologů Dont Have.
Vývoj ploutvových paprsků u některých ryb, počet zubů u zubatých velryb a vývoj srsti u savců, který je homologní s embryonálním anlagem šupin ryb a plazů, se děje prostřednictvím archalaxy.
Prostřednictvím archalaxie tak mohou v evoluci vzniknout nové orgány. Archalaxis lze pozorovat pouze ve vývoji částí organismu, nikoli však organismu jako celku. K redukci orgánů dochází také prostřednictvím fylembryogeneze. Severtsov identifikoval dva hlavní typy redukce orgánů: rudimentaci a afanezii.
✓ Rudimentace je pomalé vymizení orgánu, který ztratil svou funkci a stal se nepotřebným. Takový orgán se v ontogenezi plně nevyvine a dlouho přetrvává ve fylogenezi. Například u jeskynního obojživelníka Protea se tvoří oči a začínají se vyvíjet jako všichni obojživelníci. Mladé larvy mají oči zcela normální. Poté se zpomalí rychlost růstu oka a zastaví se vývoj extraokulárních svalů. U dospělého zvířete jsou oči malé, nehybné, zbytkové, umístěné pod epidermis.
✓ Afánizie je orgán, který je vyvinutý a funguje normálně u předků, ale u potomků se ukazuje jako škodlivý. V tomto případě může dojít k vytvoření orgánu během ontogeneze, ale poté je jeho anlage zcela resorbována. Takto je u pulců redukován ocas. Po celou dobu vývoje larev tento orgán roste a vyvíjí se. Dospělým žabám, které se pohybují skákáním, je ocas škodlivý, proto se při metamorfóze zmenší a úplně zmizí.
E. Haeckel ukázal, že změny v ontogenezi v procesu evoluce mohou vznikat prostřednictvím heterochronií – posunu v době vzniku určitého orgánu nebo struktury, a heterotopií – topografickým posunem v místě, kde se struktura vytvořila. Příkladem adaptivních heterochronií je časový posun ve formování životně důležitých orgánů u savců a lidí. V nich diferenciace předního mozku výrazně posouvá vývoj jeho ostatních částí. Jako příklady heterotopií lze uvést změnu umístění plic a plaveckého měchýře, která primárně vznikla z výrůstků ležících po stranách střeva; u potomků se plíce přesunuly na břišní stranu a plavecký měchýř na dorzální stranu střeva, příkladem je testikulární heterotopie (anabolie), pohyb srdce v strunatcích ve fylogenezi.
Díky koenogenezi, fylembryogenezi, heterotopii a heterochronii ontogeneze nejen krátce opakuje evoluční cestu, kterou prošli předci, ale také dláždí nové směry fylogeneze do budoucna.
Průzkumníci počátku 19. století. poprvé začali věnovat pozornost podobnosti fází vývoje embryí vyšších živočichů s fázemi složitosti organizace vedoucí od nízko organizovaných forem k progresivním. V roce 1828 Karl von Baer formuloval vzorec nazvaný Baerův zákon: „Čím dřívější fáze individuálního vývoje jsou porovnávány, tím více podobností lze nalézt. Při porovnání fází vývoje embryí různých druhů a tříd strunatců učinil K. Baer následující závěry.
1. Embrya zvířat stejného typu jsou v raných fázích vývoje podobná.
2. Důsledně se pohybují ve svém vývoji od obecnějších charakteristik typu ke stále specifičtějším. Nakonec se vyvinou znaky, které naznačují, že embryo patří k určitému rodu, druhu a nakonec k jednotlivým znakům.
3. Postupně se od sebe oddělují embrya různých zástupců stejného typu (obr. 1).
Obrázek 1. Podobnost embryí různých tříd obratlovců v různých stádiích
K. Baer, který nebyl evolucionistou, nedokázal spojit vzorce individuálního vývoje, které objevil, s procesem fylogeneze. Proto zobecnění, která učinil, měla význam pouze empirických pravidel.
Následný vývoj evoluční myšlenky umožnil vysvětlit podobnost raných embryí jejich historickou příbuzností a jejich osvojováním stále specifičtějších znaků s postupnou vzájemnou izolací - skutečnou izolací odpovídajících tříd, řádů, rodin, rody a druhy v procesu evoluce.
Evoluční teorie vyvinutá Charlesem Darwinem jasně zdůraznila zásadní význam problému ontogenetického vývoje. Embryonální podobnost je nyní vysvětlována skutečným vztahem organismů a jejich postupná divergence (embryonální divergence) slouží jako zřejmý odraz historické divergence těchto forem (fylogenetická divergence). V zárodku našich potomků, napsal Darwin, vidíme „nejasný portrét“ našich předků. V důsledku toho lze historii daného druhu sledovat individuálním vývojem.
Biogenetický zákon E. Haeckela
F. Müller ve svém díle „For Darwin“ (1864) formuloval myšlenku, že změny v ontogenetickém vývoji, které jsou základem procesu evoluce, mohou být vyjádřeny změnami v raných nebo pozdních fázích vývoje orgánů. V prvním případě je zachována pouze obecná podobnost mladých embryí. Ve druhém případě dochází k prodloužení a komplikaci ontogeneze spojené s přidáváním etap a opakováním (rekapitulací) v individuálním vývoji vlastností vzdálenějších dospělých předků. Müllerova práce posloužila jako základ pro formulaci základního biogenetického zákona E. Haeckela (1866), podle kterého je ontogeneze krátké a rychlé opakování fylogeneze. To znamená, že organický jedinec opakuje během rychlého a krátkého průběhu svého individuálního vývoje nejdůležitější z těch změn formy, kterými prošli jeho předkové během pomalého a dlouhého průběhu svého paleontologického vývoje podle zákonů dědičnosti a proměnlivosti. . Charakteristiky dospělých předků, které se opakují v embryogenezi jejich potomků, nazval palingenezí. Adaptace na embryonální nebo larvální stádia se nazývají cenogeneze.
Haeckelovy představy se však značně lišily od Müllerových názorů na problematiku vztahu ontogeneze a fylogeneze v procesu evoluce. Müller věřil, že evolučně nové formy vznikají změnou průběhu individuálního vývoje charakteristického pro jejich předky, tzn. změny ontogeneze jsou primární ve vztahu k fylogenetickým změnám. Fylogenetické změny podle Haeckela naopak předcházejí změnám v individuálním vývoji. Evoluční nové znaky nevznikají během ontogeneze, ale v dospělém organismu. Dospělý organismus se vyvíjí a v procesu tohoto vývoje se vlastnosti posouvají do dřívějších fází ontogeneze.
Vznikl tak problém vztahu ontogeneze a fylogeneze, který není dosud vyřešen.
Haeckel se na rozdíl od Müllera domníval, že fylogeneze je ovlivněna pouze rozšířením ontogeneze přidáním stádií a všechna ostatní stádia zůstávají nezměněna. Haeckel proto přijal pouze druhou cestu historických změn ontogeneze (podle Müllera) a jako základ fylogenetických přeměn ponechal stranou samotnou změnu fází ontogeneze. Darwin a Muller zdůrazňovali právě tuto formu vzájemné závislosti mezi ontogenezí a fylogenezí. Výklad biogenetického zákona, jak jej chápal Muller, později rozvinul A.N. Severtsov (1910-1939) v teorii fylembryogeneze. Severtsov sdílel Mullerovy názory na primát ontogenetických změn ve vztahu ke změnám v dospělých organismech a považoval ontogenezi nejen za výsledek fylogeneze, ale i za její základ. Ontogeneze se nejen prodlužuje přidáním fází: je zcela přestavěna v procesu evoluce; má svou historii, přirozeně spojenou s historií dospělého organismu a částečně ji určující.
Fylembryogeneze jsou embryonální změny spojené s fylogenetickým vývojem dospělého organismu. V procesu evoluce se všechny fáze vývoje přeskupují. Nové změny se často objevují v posledních fázích formace. Ontogeneze se komplikuje přidáváním nebo přidáváním stadií (anabolismus). Pouze v tomto případě jsou všechny předpoklady pro opakování v ontogenezi historických etap vývoje těchto částí u vzdálených předků (rekapitulace). Ontogeneze se však může změnit v jakýchkoli dalších fázích vývoje, přičemž se všechna pozdější stádia odchýlí od předchozí cesty (deviace). Konečně je možná i změna základů orgánů nebo samotných částí (archallaxe). Pak se ukazuje, že celá ontogeneze je změněna a v individuálním vývoji potomků není žádný náznak sledu průběhu historických etap vývoje jejich předků.
Biologická podstata biogenetického zákona E. Haeckela
Ve vývoji morfologie a samotné doktríny evoluce hraje důležitou roli Haeckelův biogenetický zákon a Severcovova teorie fylembryogeneze. Studium individuálního vývoje zvířat poskytlo dostatečné důkazy o jejich historickém vývoji. Biogenetický zákon je důležitou součástí metody trojitého paralelismu vyvinutého E. Haeckelem, pomocí které je rekonstruována fylogeneze. Tato metoda je založena na srovnání morfologických, embryologických a paleontologických dat. Při rekonstrukci fylogeneze morfologové stále používají Haeckeliánský princip, podle kterého ontogeneze potomků stručně opakuje a rekapituluje stadia fylogeneze předků.. Spoléháme-li se pouze na základní biogenetický zákon, nelze vysvětlit proces evoluce: nekonečný opakování toho, co bylo předáno, samo o sobě nerodí něco nového. Jelikož život na Zemi existuje díky střídání generací konkrétních organismů, dochází k jeho evoluci díky změnám v jejich ontogenezích. Tyto změny se scvrkávaly na skutečnost, že specifické ontogeneze se odchylují od cesty vytyčené rodovými formami a získávají nové rysy.
Mezi takové odchylky patří například koenogeneze – adaptace, které vznikají u embryí nebo larev a přizpůsobují je vlastnostem jejich prostředí. U dospělých organismů není cenogeneze zachována. Příkladem koenogeneze jsou rohovinové útvary v tlamě larev bezocasých obojživelníků, které jim usnadňují potravu rostlinnou potravou. Během procesu metamorfózy v žábě mizí a trávicí systém je přebudován tak, aby se živil hmyzem a červy. K cenogenezi u placentárních savců a člověka - placenta s pupeční šňůrou.
Cenogeneze, objevující se pouze v raných fázích ontogeneze, nemění typ organizace dospělého organismu, ale poskytuje vyšší pravděpodobnost přežití potomstva. Mohou být doprovázeny poklesem plodnosti a prodloužením embryonálního nebo larválního období, díky čemuž se organismus v postembryonálním nebo postlarválním období vývoje ukazuje jako zralejší a aktivnější. Poté, co vznikly a ukázaly se být užitečné, budou cenogeneze reprodukovány v následujících generacích.
Dalším typem fylogeneticky významných přeměn fylogeneze je fylembryogeneze. Představují odchylky od ontogeneze charakteristické pro předky, projevující se v embryogenezi, ale mající adaptační význam u dospělých forem. Vlasové útvary se tedy objevují u savců ve velmi raných stádiích embryonálního vývoje, ale samotný vlas je důležitý pouze u dospělých organismů.Takové změny v ontogenezi, které jsou užitečné, jsou fixovány přirozeným výběrem a reprodukovány v následujících generacích. Tyto změny jsou založeny na stejných mechanismech, které způsobují vrozené vývojové vady: narušení proliferace buněk, jejich pohyb, adheze, smrt nebo diferenciace, ale stejně jako cenogeneze se od defektů odlišují svou adaptivní hodnotou, tzn. užitečnost a fixace přirozeným výběrem ve fylogenezi.
V závislosti na fázích embryogeneze a morfogeneze specifických struktur dochází k vývojovým změnám, které mají význam fylembryogeneze, rozlišují se tři typy.
1. Anabolismus nebo extenze nastávají poté, co orgán prakticky dokončil svůj vývoj, a jsou vyjádřeny přidáním dalších fází, které mění konečný výsledek. Mezi anabolismy patří takové jevy, jako je získání specifického tvaru těla u platýse až poté, co se z vajíčka vylíhne plůdek, k nerozeznání od jiných ryb, dále vzhled zakřivení páteře, splynutí švů v mozkové lebce a konečná redistribuce krevních cév v těle savců a lidí.
2. Odchylky - odchylky, které vznikají v procesu orgánové morfogeneze. Příkladem je vývoj srdce v ontogenezi savců, kdy rekapituluje trubicové stadium, dvoukomorovou a tříkomorovou stavbu, ale stadium tvorby neúplné přepážky, charakteristické pro plazy, je nahrazeno vývojem. přepážky, konstruované a umístěné odlišně a charakteristické pouze pro savce. Ve vývoji plic u savců se nachází i rekapitulace raných stádií předků a pozdější morfogeneze probíhá novým způsobem.
3. Archalaxis - změny, které jsou detekovány na úrovni rudimentů a jsou vyjádřeny porušením jejich dělení, ranou diferenciací nebo výskytem zásadně nových analáží. Klasickým příkladem archalaxie je vývoj srsti u savců, k jejichž tvorbě dochází ve velmi raných stádiích vývoje a od samého počátku se liší od tvorby jiných kožních přívěsků obratlovců. Podle typu archalaxis se u primitivních bezlebek objevuje notochord, u chrupavčitých ryb chrupavčitá páteř a u plazů se vyvíjejí nefrony sekundární ledviny.
Je zřejmé, že během evoluce vlivem anabolismu v ontogenezích potomků se plně realizuje základní biogenetický zákon, tzn. dochází k rekapitulacím všech vývojových stádií předků. V případě odchylek se rekapitulují raná stadia předků a pozdější jsou nahrazena vývojem novým směrem. Archalaxis zcela brání rekapitulaci ve vývoji těchto struktur a mění jejich samotné základy.
V evoluci ontogeneze se anabolismus nejčastěji vyskytuje jako fylembryogeneze, která jen v malé míře mění celý vývojový proces. Odchylky jako porušení morfogenetického procesu v embryogenezi jsou přirozeným výběrem často odmítány, a jsou proto mnohem méně časté. Archallaxe se v evoluci vyskytují jen zřídka, protože mění celý průběh embryogeneze, a pokud takové změny ovlivňují základy životně důležitých orgánů nebo orgánů, které jsou důležitými embryonálními organizačními centry, pak se často ukáží jako neslučitelné se životem.
Kromě cenogeneze a fylembryogeneze lze v evoluci ontogeneze nalézt i odchylky v době vzniku orgánů - heterochronie - a místě jejich vývoje - heterotopie. První i druhý vedou ke změnám ve vzájemné korespondenci vyvíjejících se struktur a podléhají přísné kontrole přírodního výběru. Zachovány jsou pouze ty heterochronie a heterotopie, které se ukáží jako užitečné. Příkladem takových adaptivních heterochronií jsou posuny v době vzniku nejvitálnějších orgánů ve skupinách vyvíjejících se podle typu arogeneze. U savců a zejména u člověka tedy diferenciace předního mozku výrazně posouvá vývoj jeho ostatních částí.
Heterotopie vedou ke vzniku nových prostorových a funkčních spojení mezi orgány a zajišťují jejich další společný vývoj. Srdce, které se nachází pod hltanem ryb, tedy zajišťuje účinný průtok krve do žaberních tepen pro výměnu plynů. Přesouvá se do retrosternální oblasti u suchozemských obratlovců, vyvíjí se a funguje v jediném komplexu s novými dýchacími orgány - plícemi, a zde především plní funkci dodávání krve do dýchacího systému pro výměnu plynů.
Heterochronie a heterotopie, v závislosti na tom, v jakých fázích embryogeneze a orgánové morfogeneze se objevují, mohou být považovány za fylembryogenezi různých typů. Typickou anabolií je heterotopie varlete u člověka z dutiny břišní přes tříselný kanál do šourku, pozorovaná na konci embryogeneze po jeho konečném vytvoření.
Cenogeneze, fylembryogeneze, stejně jako heterotopie a heterochronie, které se ukázaly být užitečné, jsou fixovány v potomstvu a reprodukovány v následujících generacích, dokud je nevytlačí nové adaptivní změny v ontogenezi a nenahradí je. Díky tomu ontogeneze nejen stručně opakuje evoluční cestu, kterou prošli naši předkové, ale také dláždí nové směry fylogeneze do budoucna.