Was sind Stomata und wo befinden sie sich? Schutz- und Sekundärzellen der Spaltöffnungen
Spaltöffnungen, die zum epidermalen Gewebesystem gehören, sind für das Leben einer Pflanze von besonderer Bedeutung. Der Aufbau der Spaltöffnungen ist so einzigartig und ihre Bedeutung so groß, dass sie gesondert betrachtet werden sollten.
Die physiologische Bedeutung des Epidermisgewebes ist zweifach und weitgehend widersprüchlich. Einerseits ist die Epidermis strukturell so angepasst, dass sie die Pflanze vor dem Austrocknen schützt, was durch den dichten Verschluss der Epidermiszellen, die Bildung einer Kutikula und relativ lange Deckhaare begünstigt wird. Andererseits muss die Epidermis jedoch Massen von Wasserdampf und verschiedenen Gasen passieren, die in einander entgegengesetzte Richtungen strömen. Der Gas- und Dampfaustausch kann unter Umständen sehr intensiv sein. Im pflanzlichen Organismus gelingt es, diesen Widerspruch mit Hilfe von Stomata aufzulösen. Die Stomata bestehen aus zwei besonders veränderten Epidermiszellen, die durch entgegengesetzte (entlang ihrer Länge) Enden miteinander verbunden und genannt werden Schließzellen. Der Interzellularraum zwischen ihnen wird genannt Spaltöffnung.
Die Schutzzellen werden so genannt, weil sie durch aktive periodische Veränderungen des Turgors ihre Form so verändern, dass sich die Spaltöffnung entweder öffnet oder schließt. Für diese Stomata-Bewegungen sind die folgenden zwei Merkmale von großer Bedeutung. Erstens enthalten Schutzzellen im Gegensatz zu anderen Zellen der Epidermis Chloroplasten, in denen im Licht Photosynthese stattfindet und Zucker gebildet wird. Die Anreicherung von Zucker als osmotisch aktiver Substanz führt zu einer Veränderung des Turgordrucks der Schließzellen im Vergleich zu anderen Zellen der Epidermis. Zweitens verdicken sich die Membranen der Schutzzellen ungleichmäßig, sodass eine Änderung des Turgordrucks eine ungleichmäßige Änderung des Volumens dieser Zellen und folglich eine Änderung ihrer Form verursacht. Die Veränderung der Form der Schließzellen führt zu einer Veränderung der Breite der Spaltöffnung. Lassen Sie uns dies anhand des folgenden Beispiels veranschaulichen. Die Abbildung zeigt eine der Arten von Spaltöffnungen zweikeimblättriger Pflanzen. Der äußerste Teil der Stomata besteht aus häutigen Vorsprüngen, die von der Kutikula gebildet werden und manchmal unbedeutend, manchmal aber ziemlich bedeutend sind. Sie begrenzen einen kleinen Raum von der Außenfläche, dessen unterer Rand der Stomatalspalt selbst ist, genannt Stomata im Vorgarten. Hinter der Stomata-Lücke im Inneren befindet sich ein weiterer kleiner Raum, der durch kleine innere Vorsprünge der Seitenwände der Schließzellen begrenzt wird Terrassenstomata. Die Terrasse öffnet sich direkt in einen großen Interzellularraum namens Lufthohlraum.
Im Licht entsteht in den Schließzellen Zucker, er entzieht benachbarten Zellen Wasser, der Turgor der Schließzellen nimmt zu und die dünnen Teile ihrer Membran dehnen sich stärker als die dicken. Daher werden die konvexen Vorsprünge, die in den Stomata-Schlitz hineinragen, flach und die Stomata öffnen sich. Verwandelt sich beispielsweise Zucker nachts in Stärke, sinkt der Turgor in den Schließzellen, dadurch werden die dünnen Abschnitte der Schale geschwächt, sie ragen aufeinander zu und die Stomata schließen sich. In verschiedenen Pflanzen kann der Mechanismus zum Schließen und Öffnen der Spaltöffnung unterschiedlich sein. Bei Gräsern und Seggen beispielsweise haben Schließzellen verbreiterte Enden und sind im mittleren Teil verengt. Die Membranen in den mittleren Teilen der Zellen sind verdickt, während an ihren erweiterten Enden dünne Zellulosemembranen verbleiben. Eine Erhöhung des Turgors führt zu einer Schwellung der Zellenden und in der Folge dazu, dass sich die geraden Mittelteile voneinander entfernen. Dies führt zur Öffnung der Spaltöffnungen.
Merkmale des Funktionsmechanismus des Stomataapparats werden sowohl durch die Form und Struktur der Schließzellen als auch durch die Beteiligung der an die Stomata angrenzenden Epidermiszellen daran geschaffen. Unterscheiden sich die unmittelbar an die Spaltöffnungen angrenzenden Zellen im Aussehen von anderen Zellen der Epidermis, werden sie als „Stomata“ bezeichnet Begleitzellen der Spaltöffnungen.
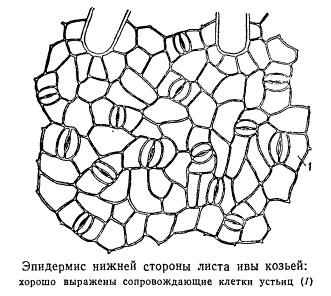
Am häufigsten haben Begleit- und Folgezellen einen gemeinsamen Ursprung.
Die Schließzellen der Spaltöffnungen sind entweder leicht über die Oberfläche der Epidermis erhaben oder umgekehrt in mehr oder weniger tiefe Gruben abgesenkt. Abhängig von der Position der Schließzellen im Verhältnis zum allgemeinen Niveau der Oberfläche der Epidermis ändert sich der eigentliche Mechanismus zur Einstellung der Breite der Spaltöffnung etwas. Manchmal verholzen die Schließzellen des Stomas, und dann wird die Regulierung der Öffnung der Stomaspalte durch die Aktivität benachbarter Epidermiszellen bestimmt. Indem sie sich ausdehnen und schrumpfen, also ihr Volumen verändern, nehmen sie die ihnen benachbarten Schließzellen mit. Stomata mit verholzten Schließzellen schließen jedoch oft überhaupt nicht. In solchen Fällen erfolgt die Regulierung der Intensität des Gas- und Dampfaustausches unterschiedlich (durch die sogenannte beginnende Trocknung). Bei Stomata mit verholzten Schließzellen bedeckt die Kutikula oft mit einer ziemlich dicken Schicht nicht nur die gesamte Spaltöffnung, sondern erstreckt sich sogar bis in die Lufthöhle und kleidet deren Boden aus.
Die meisten Pflanzen haben Stomata auf beiden Seiten des Blattes oder nur auf der Unterseite. Es gibt aber auch Pflanzen, bei denen Spaltöffnungen nur auf der Blattoberseite (bei auf der Wasseroberfläche schwimmenden Blättern) ausgebildet sind. In der Regel sind die Spaltöffnungen an den Blättern größer als an den grünen Stängeln.
Die Anzahl der Spaltöffnungen auf den Blättern verschiedener Pflanzen variiert stark. Beispielsweise beträgt die Anzahl der Spaltöffnungen auf der Unterseite eines grannenlosen Brombeerblattes durchschnittlich 30 pro 1 mm 2 , bei unter gleichen Bedingungen wachsenden Sonnenblumen sind es etwa 250. Einige Pflanzen haben bis zu 1300 Spaltöffnungen pro 1 mm 2 .
Bei Exemplaren derselben Pflanzenart hängen Dichte und Größe der Spaltöffnungen stark von den Umweltbedingungen ab. Beispielsweise befanden sich auf den Blättern einer Sonnenblume, die bei vollem Licht wuchs, durchschnittlich 220 Spaltöffnungen pro 1 mm 2 Blattoberfläche, und auf einem Exemplar, das neben der ersten, jedoch mit leichter Schattierung, wuchs, waren es etwa 140. Auf Bei einer Pflanze, die in vollem Licht wächst, nimmt die Dichte der Spaltöffnungen von den unteren Blättern zu den oberen zu.
Anzahl und Größe der Stomata hängen nicht nur stark von den Wachstumsbedingungen der Pflanze ab, sondern auch von den inneren Zusammenhängen der Lebensprozessen in der Pflanze selbst. Diese Werte (Koeffizienten) sind die empfindlichsten Reagenzien für jede Kombination von Faktoren, die das Wachstum einer Pflanze bestimmen. Daher gibt die Bestimmung der Dichte und Größe der Stomata von Blättern von Pflanzen, die unter verschiedenen Bedingungen gezüchtet wurden, eine Vorstellung von der Art der Beziehung jeder Pflanze zu ihrer Umgebung. Alle Methoden zur Bestimmung der Größe und Anzahl anatomischer Elemente in einem bestimmten Organ gehören zur Kategorie der quantitativen anatomischen Methoden, die manchmal in Umweltstudien sowie zur Charakterisierung von Kulturpflanzensorten verwendet werden, da jede Sorte einer Kulturpflanze vorhanden ist gekennzeichnet durch bestimmte Grenzen der Größe und Anzahl anatomischer Elemente pro Flächeneinheit. Methoden der quantitativen Anatomie können sowohl im Pflanzenbau als auch in der Ökologie mit großem Nutzen eingesetzt werden.
Neben Spaltöffnungen, die dem Gas- und Dampfaustausch dienen, gibt es auch Spaltöffnungen, durch die Wasser nicht in Form von Dampf, sondern in tropfenförmigem flüssigem Zustand abgegeben wird. Manchmal sind solche Stomata gewöhnlichen Stomata recht ähnlich, nur geringfügig größer, und ihre Schließzellen sind nicht beweglich. Sehr oft fehlen in solchen Stomata im voll ausgereiften Zustand Schutzzellen und es verbleibt nur ein Loch, durch das Wasser abfließt. Als Spaltöffnungen werden Stomata bezeichnet, die Tröpfchen flüssigen Wassers absondern Wasser, und alle Formationen, die an der Freisetzung von tröpfchenflüssigem Wasser beteiligt sind - Hydathoden.
Der Aufbau von Hydathoden ist vielfältig. Einige Hydathoden haben Parenchym unter dem Loch, das Wasser entfernt, das an der Übertragung von Wasser aus dem wasserleitenden System und an seiner Freisetzung aus dem Organ beteiligt ist; Bei anderen Hydathoden nähert sich das wasserführende System direkt dem Auslass. Besonders häufig bilden sich Hydathoden auf den ersten Blättern von Sämlingen verschiedener Pflanzen. So geben die jungen Blätter von Getreide, Erbsen und vielen Wiesengräsern bei feuchtwarmem Wetter tropfenweise Wasser ab. Dieses Phänomen kann in der ersten Sommerhälfte an jedem schönen Tag am frühen Morgen beobachtet werden.
Die am deutlichsten ausgeprägten Hydathoden befinden sich an den Blatträndern. Oftmals werden von jedem Zähnchen, das die Blattränder abtrennt, eine oder mehrere Hydathoden getragen.
Wenn Sie einen Fehler finden, markieren Sie bitte einen Textabschnitt und klicken Sie Strg+Eingabetaste.
Stomata einer Pflanze sind Poren, die sich in den Schichten der Epidermis befinden. Sie dienen der Verdunstung von überschüssigem Wasser und dem Gasaustausch zwischen der Blüte und der Umgebung.
Sie wurden erstmals 1675 bekannt, als der Naturforscher Marcello Malpighi seine Entdeckung in Anatome plantarum veröffentlichte. Es gelang ihm jedoch nicht, ihren wahren Zweck zu entschlüsseln, was als Anstoß für die Entwicklung weiterer Hypothesen und Forschungen diente.
Geschichte der Studie
Das 19. Jahrhundert brachte den lang erwarteten Fortschritt in der Forschung. Dank Hugo von Mohl und Simon Schwendener wurde das Grundprinzip der Funktionsweise von Spaltöffnungen und ihre Einteilung nach Strukturtyp bekannt.
Diese Entdeckungen gaben einen starken Impuls für das Verständnis der Funktionsweise von Poren, doch einige Aspekte früherer Forschungen werden bis heute weiter untersucht.
Blattstruktur
Pflanzenteile wie die Epidermis und die Stomata haben mit der inneren Struktur des Blattes zu tun, aber zuerst sollten Sie seine äußere Struktur studieren. Das Blatt besteht also aus:
- Blattspreite – ein flacher und flexibler Teil, der für Photosynthese, Gasaustausch, Wasserverdunstung und vegetative Vermehrung (bei bestimmten Arten) verantwortlich ist.
- Die Basis, in der sich die Wachstumsfuge und der Blattstiel befinden. Es hilft auch, das Blatt am Stiel zu befestigen.
- Nebenblätter sind paarige Formationen an der Basis, die die Achselknospen schützen.
- Blattstiel – der sich verjüngende Teil des Blattes, der die Blattspreite mit dem Stiel verbindet. Es ist für lebenswichtige Funktionen verantwortlich: Orientierung zum Licht und Wachstum durch Bildungsgewebe.
Die äußere Struktur des Blattes kann je nach Form und Art (einfach/komplex) leicht variieren, alle oben genannten Teile sind jedoch immer vorhanden.
Die innere Struktur umfasst die Epidermis und Stomata sowie verschiedene prägende Gewebe und Venen. Jedes der Elemente hat sein eigenes Design.
Beispielsweise besteht die Außenseite eines Blattes aus lebenden Zellen, die sich in Größe und Form unterscheiden. Die oberflächlichsten von ihnen sind transparent, sodass das Sonnenlicht in das Blatt eindringen kann.
Kleinere, etwas tiefer liegende Zellen enthalten Chloroplasten, die den Blättern ihre grüne Farbe verleihen. Aufgrund ihrer Eigenschaften wurden sie als Verschluss bezeichnet. Je nach Feuchtigkeitsgrad schrumpfen sie oder bilden untereinander Spaltöffnungen.

Struktur
Die Länge der Spaltöffnungen einer Pflanze variiert je nach Art und Grad des Lichteinfalls. Die größten Poren können eine Größe von 1 cm erreichen. Die Spaltöffnungen bilden Schutzzellen, die den Grad ihrer Öffnung regulieren.
Der Mechanismus ihrer Bewegung ist recht komplex und variiert je nach Pflanzenart. In den meisten von ihnen kann – je nach Wasserversorgung und Chloroplastengehalt – der Turgor des Zellgewebes entweder abnehmen oder zunehmen und so die Öffnung der Spaltöffnungen regulieren.

Zweck der Spaltöffnung
Auf einen Aspekt wie die Funktionen des Blattes muss wahrscheinlich nicht näher eingegangen werden. Sogar ein Schüler weiß davon. Doch wofür sind Spaltöffnungen verantwortlich? Ihre Aufgabe besteht darin, die Transpiration (den Prozess der Wasserbewegung durch eine Pflanze und ihrer Verdunstung durch äußere Organe wie Blätter, Stängel und Blüten) sicherzustellen, was durch die Arbeit von Schließzellen erreicht wird. Dieser Mechanismus schützt die Pflanze vor dem Austrocknen bei heißem Wetter und verhindert den Beginn des Fäulnisprozesses bei übermäßiger Luftfeuchtigkeit. Das Funktionsprinzip ist äußerst einfach: Wenn die Flüssigkeitsmenge in den Zellen nicht hoch genug ist, sinkt der Druck auf die Wände und die Spaltöffnung schließt sich, wodurch der lebensnotwendige Feuchtigkeitsgehalt erhalten bleibt.
Und im Gegenteil, sein Überschuss führt zu einem erhöhten Druck und der Öffnung von Poren, durch die überschüssige Feuchtigkeit verdunstet. Aus diesem Grund spielen Spaltöffnungen auch eine große Rolle bei der Kühlung von Pflanzen, da die Lufttemperatur um sie herum gerade durch Transpiration abnimmt.
Unter dem Spalt befindet sich außerdem ein Lufthohlraum, der dem Gasaustausch dient. Durch die Poren dringt Luft in die Pflanze ein und gelangt anschließend in die Atmung. Über denselben Stomataspalt entweicht dann überschüssiger Sauerstoff in die Atmosphäre. Darüber hinaus wird das Vorhandensein oder Fehlen häufig zur Klassifizierung von Pflanzen herangezogen.

Arbeitsblattfunktionen
Das Blatt ist ein äußeres Organ, über das Photosynthese, Atmung, Transpiration, Guttation und vegetative Vermehrung erfolgen. Darüber hinaus ist es in der Lage, Feuchtigkeit und organische Stoffe über die Stomata anzusammeln und der Pflanze eine bessere Anpassungsfähigkeit an schwierige Umweltbedingungen zu verleihen.
Da Wasser das wichtigste intrazelluläre Medium ist, sind die Ausscheidung und Zirkulation von Flüssigkeit im Inneren eines Baumes oder einer Blume für deren Leben gleichermaßen wichtig. In diesem Fall nimmt die Pflanze nur 0,2 % der durch sie hindurchströmenden Feuchtigkeit auf, der Rest geht an Transpiration und Guttation, wodurch die Bewegung gelöster Mineralsalze und die Abkühlung erfolgen.
Die vegetative Vermehrung erfolgt häufig durch Schneiden und Wurzeln von Blütenblättern. Viele Zimmerpflanzen werden auf diese Weise angebaut, da nur so die Reinheit der Sorte erhalten bleibt.
Wie bereits erwähnt, tragen sie zur Anpassung an verschiedene natürliche Bedingungen bei. Beispielsweise hilft die Umwandlung in Stacheln Wüstenpflanzen, die Feuchtigkeitsverdunstung zu reduzieren, Ranken verbessern die Funktionen des Stängels und große Größen dienen oft dazu, Flüssigkeit und Nährstoffe zu bewahren, wenn die klimatischen Bedingungen eine regelmäßige Auffüllung der Reserven nicht zulassen.
Und diese Liste lässt sich endlos fortsetzen. Gleichzeitig ist es schwer zu übersehen, dass diese Funktionen für die Blätter von Blumen und Bäumen gleich sind.

Welche Pflanzen haben keine Spaltöffnungen?
Da die Stomatalspalte für höhere Pflanzen charakteristisch ist, ist sie bei allen Arten vorhanden, und es ist ein Fehler, sie als nicht vorhanden zu betrachten, selbst wenn ein Baum oder eine Blume keine Blätter hat. Die einzige Ausnahme von der Regel bilden Seetang und andere Algen.
Der Aufbau der Spaltöffnungen und ihre Funktion unterscheiden sich bei Nadelbäumen, Farnen, Schachtelhalmen und Stomata von denen bei Blütenpflanzen. In den meisten von ihnen sind die Schlitze tagsüber geöffnet und nehmen aktiv am Gasaustausch und der Transpiration teil; Eine Ausnahme bilden Kakteen und Sukkulenten, deren Poren sich nachts öffnen und morgens schließen, um in trockenen Regionen Feuchtigkeit zu sparen.
Spaltöffnungen einer Pflanze, deren Blätter auf der Wasseroberfläche schwimmen, befinden sich nur in der oberen Schicht der Epidermis und bei „sitzenden“ Blättern in der unteren Schicht. Bei anderen Varianten sind diese Schlitze auf beiden Seiten der Platte vorhanden.
Lage der Stomata
Die Spaltöffnungen befinden sich auf beiden Seiten der Blattspreite, ihre Anzahl ist jedoch im unteren Teil etwas größer als im oberen Teil. Dieser Unterschied ist auf die Notwendigkeit zurückzuführen, die Feuchtigkeitsverdunstung von einer gut beleuchteten Plattenoberfläche zu reduzieren.
Bei einkeimblättrigen Pflanzen gibt es keine Besonderheiten hinsichtlich der Lage der Spaltöffnungen, da diese von der Wachstumsrichtung der Platten abhängt. Beispielsweise enthält die Epidermis vertikal ausgerichteter Pflanzenblätter sowohl in der oberen als auch in der unteren Schicht die gleiche Anzahl von Poren.

Wie bereits erwähnt, haben Schwimmblätter keine Stomata-Schlitze an der Unterseite, da sie wie reine Wasserpflanzen, die überhaupt keine solchen Poren haben, Feuchtigkeit über die Kutikula aufnehmen.
Die Spaltöffnungen von Nadelbäumen liegen tief unter der Endodermis, was zu einer verminderten Transpirationsfähigkeit beiträgt.
Außerdem unterscheidet sich die Lage der Poren relativ zur Oberfläche der Epidermis. Die Schlitze können auf gleicher Höhe mit den übrigen „Hautzellen“ liegen, höher oder tiefer verlaufen, regelmäßige Reihen bilden oder willkürlich über das Hautgewebe verteilt sein.
Bei Kakteen, Sukkulenten und anderen Pflanzen, deren Blätter fehlen oder sich verändert haben und sich in Nadeln verwandelt haben, befinden sich Spaltöffnungen an den Stängeln und fleischigen Teilen.
Typen
Die Spaltöffnungen einer Pflanze werden je nach Lage der begleitenden Zellen in viele Arten eingeteilt:
- Anomozytär – gilt als die häufigste Form, bei der sich die Nebenprodukte nicht von anderen in der Epidermis vorkommenden unterscheiden. Eine seiner einfachen Modifikationen kann als Laterozytentyp bezeichnet werden.
- Parazytisch – gekennzeichnet durch parallele Aneinanderreihung von Begleitzellen relativ zur Spaltöffnung.
- Diazit – hat nur zwei Seitenteilchen.
- Anisozytär – ein Typ, der nur in Blütenpflanzen vorkommt, mit drei Begleitzellen, von denen eine deutlich unterschiedlich groß ist.
- Tetrazytär – charakteristisch für Monokotyledonen, hat vier Begleitzellen.
- Enzyklozytisch – darin schließen sich die Seitenteilchen ringförmig um die schließenden.
- Perizytisch – es zeichnet sich durch Stomata aus, die nicht mit der Begleitzelle verbunden sind.
- Desmozyten – unterscheidet sich vom vorherigen Typ nur durch das Vorhandensein einer Adhäsion zwischen der Lücke und dem Seitenpartikel.
Hier sind nur die beliebtesten Typen aufgeführt.
Der Einfluss von Umweltfaktoren auf die äußere Struktur des Blattes
Für das Überleben einer Pflanze ist der Grad ihrer Anpassungsfähigkeit äußerst wichtig. Beispielsweise zeichnen sich feuchte Gebiete durch große Blattspreiten und eine große Anzahl von Stomata aus, während dieser Mechanismus in trockenen Regionen anders funktioniert. Weder Blumen noch Bäume unterscheiden sich in der Größe und die Anzahl der Poren ist deutlich reduziert, um eine übermäßige Verdunstung zu verhindern.

So lässt sich nachvollziehen, wie sich Pflanzenteile im Laufe der Zeit unter dem Einfluss der Umwelt verändern, was sich auch auf die Anzahl der Spaltöffnungen auswirkt.
Bestimmung des Stomata-Zustands bei Zimmerpflanzen
Das Blatt einer Pflanze erfüllt verschiedene Funktionen. Dies ist das Hauptorgan, in dem Photosynthese, Gasaustausch und Transpiration (Verdunstung von Wasser) stattfinden. Um den Gasaustausch durchzuführen, verfügen die Landorgane der Pflanze über spezielle Formationen – Spaltöffnungen.
Obwohl Stomata Teil der Epidermis (Blatthaut) sind, handelt es sich um besondere Zellgruppen. Der Stomatalapparat besteht aus zwei Schließzellen, zwischen denen sich eine Stomatalspalte, 2–4 parastomatäre Zellen und eine unter der Stomatalspalte befindliche Gas-Luft-Kammer befinden.
Die Schließzellen der Stomata haben eine längliche, gebogene, „bohnenförmige“ Form. Ihre der Stomatalspalte zugewandten Wände sind verdickt. Stomatalzellen können ihre Form ändern – dadurch öffnet oder schließt sich die Stomatalspalte. Diese Zellen enthalten Chloroplasten (grüne Plastiden). Das Öffnen und Schließen der Spaltöffnung erfolgt aufgrund von Änderungen des Turgors (osmotischer Druck) in den Schließzellen. Die Chloroplasten der Schließzellen enthalten Stärke, die in Zucker umgewandelt werden kann. Bei der Umwandlung von Stärke in Zucker steigt der osmotische Druck und die Spaltöffnungen öffnen sich. Bei sinkendem Zuckergehalt erfolgt der umgekehrte Vorgang und die Spaltöffnungen schließen sich.
Spaltöffnungen sind am frühen Morgen oft weit geöffnet und tagsüber geschlossen (oder halbgeschlossen). Die Anzahl der Stomata hängt von den Umgebungsbedingungen (Temperatur, Licht, Luftfeuchtigkeit) ab. Der Grad ihrer Öffnung zu verschiedenen Tageszeiten variiert je nach Art stark. In den Blättern von Pflanzen in feuchten Lebensräumen beträgt die Spaltöffnungsdichte 100–700 pro 1 mm2.
Bei den meisten Landpflanzen befinden sich Spaltöffnungen nur auf der Blattunterseite. Sie können sich auch auf beiden Seiten des Blattes befinden, wie zum Beispiel bei Kohl oder Sonnenblume. Darüber hinaus ist die Dichte der Stomata auf der Ober- und Unterseite des Blattes nicht gleich: Bei Kohl beträgt sie 140 bzw. 240 pro 1 mm2 und bei Sonnenblume 175 bzw. 325 pro 1 mm2. Bei Wasserpflanzen wie Seerosen befinden sich Stomata nur auf der Blattoberseite mit einer Dichte von etwa 500 pro 1 mm 2. Unterwasserpflanzen haben überhaupt keine Spaltöffnungen.
Ziel der Arbeit:
Bestimmung des Stomata-Zustands bei verschiedenen Zimmerpflanzen.
Aufgaben
1. Studieren Sie die Frage nach Struktur, Lage und Anzahl der Stomata in verschiedenen Pflanzen anhand zusätzlicher Literatur.
2. Wählen Sie Pflanzen für die Forschung aus.
3. Bestimmen Sie den Zustand der Stomata und den Grad ihrer Öffnung in verschiedenen Zimmerpflanzen, die im Biologieunterricht verfügbar sind.
Materialen und Methoden
Die Bestimmung des Stomata-Zustands erfolgte nach der in „Methodische Empfehlungen für die Pflanzenphysiologie“ (zusammengestellt von E.F. Kim und E.N. Grishina) beschriebenen Methode. Der Kern der Technik besteht darin, dass der Grad der Öffnung der Spaltöffnungen durch das Eindringen bestimmter Chemikalien in das Fruchtfleisch des Blattes bestimmt wird. Zu diesem Zweck werden verschiedene Flüssigkeiten verwendet: Äther, Alkohol, Benzin, Kerosin, Benzol, Xylol. Wir verwendeten Alkohol, Benzol und Xylol, die uns im Chemieraum zur Verfügung gestellt wurden. Das Eindringen dieser Flüssigkeiten in das Fruchtfleisch des Blattes hängt vom Öffnungsgrad der Spaltöffnungen ab. Wenn 2–3 Minuten nach dem Auftragen eines Flüssigkeitstropfens auf die Unterseite der Blattspreite ein heller Fleck auf dem Blatt erscheint, bedeutet dies, dass die Flüssigkeit durch die Spaltöffnungen eindringt. In diesem Fall dringt Alkohol nur bei weit geöffneten Spaltöffnungen in das Blatt ein, Benzol bereits mit einer durchschnittlichen Öffnungsweite und nur Xylol dringt durch fast geschlossene Spaltöffnungen ein.
Im ersten Arbeitsschritt haben wir versucht, die Möglichkeit zu etablieren, den Zustand der Spaltöffnungen (Öffnungsgrad) verschiedener Pflanzen zu bestimmen. In diesem Experiment wurden Agave, Cyperus, Tradescantia, Geranie, Oxalis, Syngonium, Amazonas-Lilie, Begonie, Sanchetia, Dieffenbachia, Clerodendron, Passionsblume, Kürbis und Bohne verwendet. Für die weitere Bearbeitung wurden Oxalis, Geranie, Begonie, Sanchetia, Clerodendron, Passionsblume, Kürbis und Bohnen ausgewählt. In anderen Fällen konnte der Grad der Spaltöffnung nicht bestimmt werden. Dies kann auf die Tatsache zurückzuführen sein, dass Agave, Cyperus und Lilie ziemlich harte Blätter haben, die mit einer Beschichtung bedeckt sind, die das Eindringen von Substanzen durch den Stomatalspalt verhindert. Ein weiterer möglicher Grund könnte sein, dass ihre Stomata zum Zeitpunkt des Experiments (14.00 Uhr) bereits geschlossen waren.
Die Studie wurde über eine Woche durchgeführt. Jeden Tag nach der Schule um 14.00 Uhr bestimmten wir mit der oben genannten Methode den Grad der Spaltöffnung.
Resultate und Diskussion
Die erhaltenen Daten sind in der Tabelle dargestellt. Die angegebenen Daten sind gemittelt, weil An verschiedenen Tagen war der Zustand der Spaltöffnungen unterschiedlich. So wurde bei sechs Messungen die weite Öffnung der Spaltöffnungen zweimal bei Waldsauerampfer, einmal bei Geranien und bei Begonien zweimal der durchschnittliche Öffnungsgrad der Spaltöffnungen gemessen. Diese Unterschiede hängen nicht vom Zeitpunkt des Experiments ab. Möglicherweise hängen sie mit den klimatischen Bedingungen zusammen, obwohl die Temperatur im Büro und die Beleuchtung der Pflanzen recht konstant waren. Daher können die erhaltenen gemittelten Daten als eine bestimmte Norm für diese Pflanzen angesehen werden.
Die Studie zeigt, dass bei verschiedenen Pflanzen zur gleichen Zeit und unter den gleichen Bedingungen der Grad der Spaltöffnung nicht gleich ist. Es gibt Pflanzen mit weit geöffneten Spaltöffnungen (Begonie, Sanchetia, Kürbis) und mittelgroßen Spaltöffnungen (Oxalis, Geranie, Bohnen). Schmale Spaltöffnungen kommen nur im Clerodendron vor.
Wir betrachten diese Ergebnisse als vorläufig. Zukünftig wollen wir herausfinden, ob und wie es biologische Rhythmen beim Öffnen und Schließen von Spaltöffnungen in verschiedenen Pflanzen gibt. Dazu wird tagsüber der Zustand der Spaltöffnungen überwacht.
Stomata, ihre Struktur und Wirkungsmechanismus
Epidermiszellen sind aufgrund der besonderen Struktur ihrer Außenwand für Wasser und Gase nahezu undurchdringlich. Wie erfolgt der Gasaustausch zwischen der Anlage und der Außenumgebung sowie die Wasserverdunstung – Prozesse, die für das normale Funktionieren der Anlage notwendig sind? Unter den Zellen der Epidermis gibt es charakteristische Formationen, die Stomata genannt werden.
Die Stomata sind eine schlitzartige Öffnung, die auf beiden Seiten von zwei meist halbmondförmigen Schließzellen begrenzt wird.
Stomata sind Poren in der Epidermis, durch die der Gasaustausch stattfindet. Man findet sie vor allem in den Blättern, aber auch am Stängel. Jedes Stomata ist auf beiden Seiten von Schließzellen umgeben, die im Gegensatz zu anderen Epidermiszellen Chloroplasten enthalten. Schließzellen steuern die Größe der Stomata-Öffnung, indem sie ihre Prallheit verändern.
Diese Zellen sind lebend und enthalten Chlorophyllkörner und Stärkekörner, die in anderen Zellen der Epidermis fehlen. Auf dem Blatt sind besonders viele Spaltöffnungen vorhanden. Der Querschnitt zeigt, dass sich direkt unter den Spaltöffnungen im Blattgewebe ein Hohlraum befindet, der Atemhöhle genannt wird. Innerhalb der Lücke liegen die Schutzzellen im mittleren Teil der Zellen näher beieinander, darüber und darunter sind sie weiter voneinander entfernt und bilden Räume, die als Vorder- und Hinterhof bezeichnet werden.
Schließzellen sind in der Lage, ihre Größe zu vergrößern und zu verkleinern, wodurch sich die Spaltöffnung manchmal weit öffnet, manchmal enger wird oder sogar vollständig geschlossen wird.
Somit sind Schließzellen der Apparat, der den Prozess des Öffnens und Schließens der Spaltöffnungen reguliert.
Wie wird dieser Prozess durchgeführt?
Die dem Spalt zugewandten Wände der Schließzellen sind viel dicker als die Wände, die benachbarten Epidermiszellen zugewandt sind. Wenn die Pflanze beleuchtet ist und überschüssige Feuchtigkeit hat, reichert sich Stärke in den Chlorophyllkörnern der Schließzellen an, die teilweise in Zucker umgewandelt wird. Im Zellsaft gelöster Zucker zieht Wasser aus benachbarten Epidermiszellen an, wodurch der Turgor in den Schließzellen zunimmt. Durch starken Druck treten die Wände der an die Epidermis angrenzenden Zellen hervor und die gegenüberliegenden, stark verdickten Wände werden begradigt. Dadurch öffnet sich die Spaltöffnung und der Gasaustausch sowie die Wasserverdunstung nehmen zu. Bei Dunkelheit oder Feuchtigkeitsmangel sinkt der Turgordruck, die Schließzellen kehren in ihre ursprüngliche Position zurück und die verdickten Wände schließen sich. Der Stomatalspalt schließt sich.
Stomata befinden sich auf allen jungen, nicht verholzten Bodenorganen der Pflanze. Besonders viele davon finden sich auf den Blättern, hier befinden sie sich hauptsächlich auf der Unterseite. Steht das Blatt senkrecht, bilden sich auf beiden Seiten Spaltöffnungen. Bei den Blättern einiger auf der Wasseroberfläche schwimmender Wasserpflanzen (z. B. Seerosen, Eikapseln) befinden sich Spaltöffnungen nur auf der Blattoberseite.
Anzahl der Spaltöffnungen pro 1 Quadratmeter. Die Blattoberfläche beträgt im Durchschnitt 300 mm, manchmal aber auch 600 oder mehr. Rohrkolben (Typha) hat über 1300 Spaltöffnungen pro Quadratmeter. mm. In Wasser getauchte Blätter haben keine Spaltöffnungen. Die Spaltöffnungen sind meist gleichmäßig über die gesamte Hautoberfläche verteilt, bei manchen Pflanzen sind sie jedoch in Gruppen gesammelt. Bei einkeimblättrigen Pflanzen sowie auf den Nadeln vieler Nadelbäume liegen sie in Längsreihen. Bei Pflanzen in Trockengebieten sind die Spaltöffnungen oft in das Blattgewebe eingetaucht. Die stomatale Entwicklung verläuft normalerweise wie folgt. In einzelnen Zellen der Epidermis bilden sich bogenförmige Wände, die die Zelle in mehrere kleinere teilen, sodass die mittlere zum Vorfahren der Stomata wird. Diese Zelle ist durch ein Längsseptum (entlang der Zellachse) geteilt. Dieses Septum spaltet sich dann und es entsteht ein Spalt. Die Zellen, die es begrenzen, werden zu den Schutzzellen der Spaltöffnungen. Einige Lebermoose haben besondere Stomata ohne Schließzellen.
In Abb. zeigt das Aussehen von Stomata und Schließzellen in einer mikroskopischen Aufnahme, die mit einem Rasterelektronenmikroskop erstellt wurde.
Hier ist zu erkennen, dass die Zellwände der Schließzellen unterschiedlich dick sind: Die Wand, die näher an der Öffnung der Stomata liegt, ist deutlich dicker als die gegenüberliegende Wand. Darüber hinaus sind die Zellulose-Mikrofibrillen, aus denen die Zellwand besteht, so angeordnet, dass die dem Loch zugewandte Wand weniger elastisch ist und einige Fasern eine Art Ring um die Schließzellen bilden, ähnlich wie bei Würstchen. Wenn die Zelle Wasser aufnimmt und prall wird, verhindern diese Ringe, dass sie sich weiter ausdehnt, sodass sie sich nur in die Länge ausdehnen kann. Da die Schließzellen an ihren Enden verbunden sind und sich die dünneren Wände abseits der Spaltöffnung leichter ausdehnen, nehmen die Zellen eine halbkreisförmige Form an. Daher entsteht zwischen den Schutzzellen ein Loch. (Den gleichen Effekt erzielen wir, wenn wir einen wurstförmigen Ballon aufblasen, an dessen einer Seite Klebeband befestigt ist.)
Wenn umgekehrt Wasser die Schließzellen verlässt, schließt sich die Pore. Wie es zu der Veränderung der Zellpralligkeit kommt, ist noch nicht klar.
Eine der traditionellen Hypothesen, die „Zucker-Stärke“-Hypothese, besagt, dass im Laufe des Tages die Zuckerkonzentration in den Schließzellen zunimmt und dadurch der osmotische Druck in den Zellen und der Wasserfluss in sie zunimmt. Bisher konnte jedoch niemand nachweisen, dass sich in den Schließzellen genügend Zucker ansammelt, um die beobachteten Veränderungen des osmotischen Drucks zu verursachen. Kürzlich wurde festgestellt, dass sich tagsüber im Licht Kaliumionen und begleitende Anionen in Schließzellen ansammeln; Diese Ansammlung von Ionen reicht völlig aus, um die beobachteten Veränderungen hervorzurufen. Im Dunkeln verlassen Kaliumionen (K+) die Schließzellen und gelangen in die angrenzenden Epidermiszellen. Es ist noch unklar, welches Anion die positive Ladung des Kaliumions ausgleicht. Einige (aber nicht alle) der untersuchten Pflanzen zeigten die Anreicherung großer Mengen an Anionen organischer Säuren wie Malat. Gleichzeitig verkleinern sich Stärkekörner, die im Dunkeln in den Chloroplasten der Schließzellen erscheinen. Dies deutet darauf hin, dass Stärke im Licht in Malat umgewandelt wird.
Einige Pflanzen, wie zum Beispiel Allium cepa (Zwiebel), haben keine Stärke in ihren Schließzellen. Wenn die Spaltöffnungen geöffnet sind, sammelt sich Malat daher nicht an und Kationen werden offenbar zusammen mit anorganischen Anionen wie Chlorid (Cl-) absorbiert.
Einige Fragen bleiben ungelöst. Warum braucht man zum Beispiel Licht, um Spaltöffnungen zu öffnen? Welche Rolle spielen Chloroplasten neben der Speicherung von Stärke? Verwandelt sich Malat im Dunkeln wieder in Stärke? Im Jahr 1979 wurde gezeigt, dass den Chloroplasten der Schließzellen von Vicia faba (Ackerbohne) Enzyme des Calvin-Zyklus fehlen und das Thylakoidsystem schlecht entwickelt ist, obwohl Chlorophyll vorhanden ist. Dadurch funktioniert der übliche C3-Weg der Photosynthese nicht und es wird keine Stärke gebildet. Dies könnte erklären, warum Stärke nicht wie in gewöhnlichen Photosynthesezellen tagsüber, sondern nachts gebildet wird. Eine weitere interessante Tatsache ist das Fehlen von Plasmodesmen in Schließzellen, d. h. vergleichende Isolierung dieser Zellen von anderen Zellen der Epidermis.
Bewegungen.
Spaltöffnungen erfüllen zwei Hauptfunktionen: Sie führen den Gasaustausch und die Transpiration (Verdunstung) durch.
Die Stomata bestehen aus zwei Schließzellen und einem Spalt dazwischen. Angrenzend an die Schließzellen befinden sich sekundäre (parostomatale) Zellen. Unter den Spaltöffnungen befindet sich ein Lufthohlraum. Die Spaltöffnungen können sich je nach Bedarf automatisch schließen oder öffnen. Dies ist auf Turgorphänomene zurückzuführen.
Der Grad der Spaltöffnung hängt von der Lichtintensität, der Wassermenge im Blatt und dem Kohlendioxid ab. in Interzellularräumen, Lufttemperatur und anderen Faktoren. Abhängig vom Faktor, der den motorischen Mechanismus auslöst (Licht oder beginnender Wassermangel im Blattgewebe), werden photo- und hydroaktive Bewegung der Stomata unterschieden. Es gibt auch hydrostatische Bewegungen, die durch Veränderungen im Wassergehalt der Epidermiszellen verursacht werden und den Stoffwechsel der Schließzellen nicht beeinflussen. Beispielsweise kann ein tiefer Wassermangel dazu führen, dass ein Blatt welkt, während die Epidermiszellen, die kleiner werden, die Schließzellen dehnen und sich die Stomata öffnen. Oder umgekehrt: Unmittelbar nach dem Regen schwellen die Epidermiszellen so stark an
aus Wasser, das die Schließzellen zusammendrückt und die Stomata schließen.
Hydropassive Lösung – Schließt Stomata-Schlitze, wenn das Zellparenchym mit Wasser überfüllt ist und komprimiert die Endzellen mechanisch
Hydroaktives Öffnen und Schließen sind Bewegungen, die durch Veränderungen im Wassergehalt der Schließzellen der Stomata verursacht werden.
Photoaktiv – manifestiert sich im Öffnen der Stomata im Licht und im Schließen im Dunkeln.
13. Einfluss äußerer Faktoren auf die Transpiration
Unter Transpiration versteht man den Feuchtigkeitsverlust in Form der Verdunstung von Wasser von der Oberfläche von Blättern oder anderen Pflanzenteilen, der mithilfe von Stomata erfolgt. Bei Wassermangel im Boden verringert sich die Transpiration.
Niedrig Temperatur inaktivieren Enzyme, erschweren die Wasseraufnahme und verlangsamen die Transpiration. Hohe Temperaturen führen zu einer Überhitzung der Blätter und einer erhöhten Transpiration. Mit steigender Temperatur erhöht sich die Transpiration. Die Temperatur ist die Energiequelle für die Wasserverdunstung. Der kühlende Effekt der Transpiration kommt vor allem bei hohen Temperaturen, geringer Luftfeuchtigkeit und guter Wasserversorgung zum Tragen. Darüber hinaus erfüllt die Temperatur auch eine regulierende Funktion und beeinflusst den Grad der Öffnung der Spaltöffnungen.
Licht. Im Licht steigt die Temperatur des Blattes und die Transpiration nimmt zu, und die physiologische Wirkung des Lichts besteht in seiner Auswirkung auf die Bewegung der Stomata – Pflanzen transpirieren im Licht stärker als im Dunkeln. Die Wirkung von Licht auf die Transpiration beruht hauptsächlich auf der Tatsache, dass grüne Zellen nicht nur Infrarot-Sonnenlicht absorbieren, sondern auch sichtbares Licht, das für die Photosynthese notwendig ist. Bei völliger Dunkelheit schließen sich die Spaltöffnungen zunächst vollständig und öffnen sich dann leicht.
Wind Erhöht die Transpiration aufgrund der Mitnahme von Wasserdampf, wodurch ein Mangel an der Oberfläche der Blätter entsteht. Die Windgeschwindigkeit beeinflusst die Transpiration nicht so stark wie die Verdunstung an der freien Wasseroberfläche. Wenn der Wind aufkommt und seine Geschwindigkeit zunimmt, nimmt die Transpiration zunächst zu, eine weitere Verstärkung des Windes hat jedoch fast keinen Einfluss auf diesen Prozess.
Luftfeuchtigkeit. Bei zu hoher Luftfeuchtigkeit nimmt die Transpiration ab (in Gewächshäusern), bei trockener Luft nimmt sie zu; je niedriger die relative Luftfeuchtigkeit der Luft, desto geringer ist ihr Wasserpotential und desto schneller erfolgt die Transpiration. Bei Wassermangel im Blatt erfolgt die stomatale und extrastomatale Regulierung aktiviert, so dass die Intensität der Transpiration langsamer zunimmt als die Verdunstung von Wasser von der Wasseroberfläche. Bei einem starken Wasserdefizit kann die Transpiration trotz zunehmender Trockenheit der Luft nahezu zum Erliegen kommen. Mit zunehmender Luftfeuchtigkeit nimmt die Transpiration ab; Bei hoher Luftfeuchtigkeit kommt es lediglich zur Guttation.
Eine hohe Luftfeuchtigkeit stört den normalen Transpirationsverlauf und wirkt sich daher negativ auf den Aufwärtstransport von Substanzen durch Gefäße, die Regulierung der Pflanzentemperatur und die Spaltöffnungsbewegungen aus.
Wassermangel ist ein Mangel an Wasser für Pflanzen.



